Marea majoritate medicamente are efect terapeutic prin schimbarea activității sistemelor fiziologice ale celulelor care sunt produse în organism în timpul evoluției. Sub influența unei substanțe medicinale în organism, de regulă, nu apare un nou tip de activitate celulară, doar viteza diferitelor procese naturale se modifică. Inhibarea sau excitarea proceselor fiziologice conduce la scăderea sau îmbunătățirea funcțiilor corespunzătoare ale țesuturilor corpului.
Medicamentele pot acționa asupra receptorilor specifici, enzimelor, membranelor celulare sau pot interacționa direct cu substanțele celulare. Detalii despre mecanismele de acțiune substanțe medicinale studiat într-un curs de farmacologie generală sau experimentală. Mai jos oferim doar câteva exemple ale principalelor mecanisme de acțiune ale medicamentelor.
Efect asupra receptorilor specifici. Receptorii sunt structuri macromoleculare sensibile selectiv la anumiți compuși chimici. Interacțiunea substanțelor chimice cu receptorul duce la apariția unor modificări biochimice și fiziologice în organism, care sunt exprimate într-un efect clinic particular.
Medicamentele care excită sau cresc direct activitatea funcțională a receptorilor se numesc agoniști, iar substanțele care inhibă acțiunea agoniștilor specifici sunt numite antagoniști. Antagonismul poate fi competitiv și necompetitiv. În primul caz, substanța medicamentoasă concurează cu un regulator natural (mediator) pentru siturile de legare în receptorii specifici. Un blocaj al receptorului cauzat de un antagonist competitiv poate fi eliminat cu doze mari de agonist sau mediator natural.
O varietate de receptori sunt împărțiți de sensibilitatea la mediatorii naturali și la antagoniștii acestora. De exemplu, receptorii sensibili la acetilcolină sunt numiți colinergici, receptorii sensibili la adrenalină sunt numiți adrenergici. În ceea ce privește sensibilitatea la muscarină și nicotină, receptorii colinergici sunt împărțiți în muscarinici sensibili (receptori m-colinergici) și nicotină-sensibili (n-colinergici). Receptorii H-colinergici sunt eterogeni. S-a stabilit că diferența lor constă în sensibilitatea la diverse substanțe. Există receptori n-colinergici localizați în ganglionii sistemului nervos autonom și receptori n-colinergici ai mușchiului striat. Sunt cunoscute diverse subtipuri de receptori adrenergici, notate cu litere grecești α1, α 2, β1, β2.
Sunt izolate și histamina H1 și H2, dopamina, serotonina, opioidul și alți receptori.
Efect asupra activității enzimelor. Unele medicamente cresc sau inhibă activitatea enzimelor specifice. De exemplu, fizostigmina și neostigmina reduc activitatea colinesterazei, care distruge acetilcolina și dau efecte caracteristice excitării sistemului nervos parasimpatic. Inhibitorii monoaminoxidazei (iprazid, nialamidă), care împiedică distrugerea adrenalinei, cresc activitatea sistemului nervos simpatic. Fenobarbitalul și zixorina, crescând activitatea glucuronil transferazei hepatice, reduc nivelul bilirubinei din sânge.
Efect fizico-chimic asupra membranelor celulare. Activitatea celulelor sistemelor nervoase și musculare depinde de fluxul de ioni care determină potențialul electric transmembranar. Unele medicamente modifică transportul ionic.
Deci medicamente antiaritmice, anticonvulsivante, medicamente pentru anestezie generală.
Interacțiune chimică directă. Medicamentele pot interacționa direct cu molecule mici sau ioni în interiorul celulelor. De exemplu, acidul etilenediaminetetraacetic (EDTA) leagă puternic ionii de plumb. Principiul interacțiunii chimice directe stă la baza utilizării multor antidoturi pentru intoxicații chimice. Un alt exemplu este neutralizarea acidului clorhidric cu antiacide.
farmacodinamie
Studiază mecanismul de acțiune al medicamentelor, precum și efectele biochimice și fiziologice ale acestora. Sarcinile ei includ o descriere a interacțiunilor chimice și fizice dintre medicament și celula țintă, precum și spectrul complet și severitatea efectelor farmacologice ale acestuia. Cunoașterea tiparelor farmacodinamice vă permite să alegeți medicația potrivită. Studiile farmacodinamice oferă o înțelegere mai profundă a reglării proceselor biochimice și fiziologice în organism (Katzung B.G., 1998; Lawrence D.R. și colab., 2002).
Acțiunea majorității medicamentelor este mediată de legarea lor la macromoleculele organismului. O schimbare a stării funcționale a acestor macromolecule, determină, la rândul său, un lanț de reacții biochimice și fiziologice care sunt transformate într-un efect farmacologic. Macromoleculele cu care interacționează substanțele chimice se numesc receptori. Astfel, orice macromolecule active funcțional pot servi ca receptori pentru medicamente. Mai multe consecințe importante decurg din această afirmație. În primul rând, cu ajutorul medicamentelor, puteți modifica viteza oricărui proces fiziologic din organism. În al doilea rând, medicamentele nu fac decât să schimbe funcțiile fiziologice naturale ale celulei, fără a-i da noi proprietăți.
receptorii
Majoritatea receptorilor sunt proteine. Aceștia sunt receptori de hormoni, factori de creștere, mediatori, proteine \u200b\u200bimplicate în cele mai importante reacții metabolice și de reglare (dihidrofolat reductază, acetilcolinesterază), proteine \u200b\u200bde transport (Na +, K + -ATPase), proteine \u200b\u200bstructurale (tubulină). Componentele celulare de natură chimică diferită, cum ar fi acizii nucleici, cu care interacționează agenții antitumorali, pot acționa, de asemenea, ca receptori.
Receptorii factorilor regulatori endogeni - hormoni, mediatori etc., au semnificație farmacologică. Acești receptori servesc drept ținte pentru multe medicamente, acționând de obicei selectiv datorită specificității ridicate a receptorilor pentru liganzi endogeni. Medicamentele care, după ce se leagă de receptor, reproduc efectul fiziologic al ligandului endogen, se numesc aganisti sau stimulente. Medicamentele care nu provoacă acest efect, dar inhibă legarea liganzilor endogeni, sunt numiți antagoniști sau blocanți. Substanțele al căror efect este mai puțin pronunțat decât efectul agoniștilor se numesc agoniști parțiali. Preparatele care stabilizează receptorul într-o conformație neactivată sunt clasificate ca agoniști inversi.
Dependență structurală și funcțională
Structura chimică a medicamentului determină destul de rigid afinitatea acestuia pentru receptori și activitatea internă. O ușoară modificare a structurii chimice poate afecta semnificativ proprietățile farmacologice.
Sinteza noilor medicamente se bazează în mare măsură pe aceasta. Deoarece modificarea chimică nu afectează în mod necesar toate proprietățile farmacologice în mod egal, este posibilă îmbunătățirea eficacității și siguranței medicamentului, creșterea selectivității sale și îmbunătățirea caracteristicilor farmacocinetice. De exemplu, mulți antagoniști hormonali și mediatori utilizați în clinică sunt sintetizați prin modificarea chimică a substanțelor endogene.
Puncte de aplicare a drogurilor
Deoarece efectul medicamentelor este mediat de receptori, punctul de aplicare al medicamentului este determinat nu numai de caracteristicile distribuției sale, ci și de localizarea receptorilor, iar efectele farmacologice depind de semnificația funcțională a acestor receptori. Efectele farmacologice ale medicamentelor ai căror receptori sunt comuni în multe organe și țesuturi sunt diverse. Dacă acești receptori îndeplinesc o funcție vitală pentru celule, nu numai că este dificil să utilizați medicamentul în scop terapeutic, dar este, de asemenea, nesigur. Cu toate acestea, aceste medicamente pot avea o importanță clinică deosebită. Deci, glicozidele cardiace, utilizate pe scară largă în insuficiența cardiacă, modifică transportul ionilor prin membrana celulară, de care depinde activitatea vitală a celulei. Au un interval terapeutic îngust și sunt foarte toxice. Un alt exemplu sunt agenții antitumorali. Dacă receptorii cu care medicamentul interacționează sunt prezenți doar pe câteva tipuri de celule diferențiate, efectul său este mai selectiv. Aceste medicamente pot avea mai puține reacții adverse, dar totuși, aceste medicamente pot fi toxice dacă receptorii lor îndeplinesc o funcție vitală. Unele otrăvuri biologice (toxina botulinică etc.) acționează într-un mod similar. În plus, chiar dacă efectul farmacologic direct este selectiv, consecințele acestuia pot fi mai diverse.
Receptori endogeni ai factorilor de reglare
Termenul de receptor se referă la orice componentă macromoleculară a unei celule de care se leagă un medicament. Unul dintre cei mai importanți receptori ai medicamentelor sunt proteinele celulare, care servesc ca receptori pentru factori de reglare endogeni - hormoni, factori de creștere, mediatori. Prin legarea la ligandul endogen, receptorii transmit semnalul din acesta în celula țintă.
De la receptor, semnalul ajunge la țintele celulare (proteine \u200b\u200befectoare) direct sau prin molecule de semnalizare intermediare - convertoare de proteine. Receptorul, convertorii de proteine \u200b\u200bși proteinele efectoare formează sistemul receptor-efector. Cea mai apropiată proteină efector din lanțul de transmitere a semnalului nu este adesea un efector terminal (care afectează direct funcțiile celulare), ci o enzimă sau proteină de transport implicată în formarea, transportul sau inactivarea unui al doilea mediator - un ion sau o moleculă mică. Al doilea mediator, la rândul său, transferă informația către o varietate de ținte intracelulare, asigurând răspunsul lor simultan la un semnal de la un receptor.
Receptorii, proteinele de conversie și proteinele efectoare nu numai că transmit informații. De asemenea, coordonează semnale de la diferiți liganzi, pe de o parte, și toate aceste semnale cu procese metabolice în celulă, pe de altă parte.
Acționând ca catalizatori, receptorii sporesc semnalul biologic. Datorită acestei proprietăți importante, ele servesc drept ținte excelente pentru medicamente. Totuși, amplificatorii de semnal nu sunt numai receptori cu activitate enzimatică, ci și toți receptorii cunoscuți. Într-adevăr, când o singură moleculă de ligand se leagă de un receptor conjugat la un canal ionic, mulți ioni trec prin acesta din urmă. Același lucru este valabil și pentru receptorii hormonilor steroizi: o moleculă hormonală declanșează transcrierea multor copii de mARN, pe baza cărora sunt sintetizate numeroase molecule de proteine.
În funcție de structura și mecanismul de acțiune, receptorii substanțelor biologic active sunt împărțiți în mai multe clase. Numărul acestor clase este mic.
Receptori enzimatici
Cel mai mare grup de receptori cu activitate enzimatică sunt receptorii cu membrană cu propria activitate de proteină kinaza. Fosforilează o varietate de proteine \u200b\u200befectoare situate pe interiorul membranei celulare. Drept urmare, funcția acestor proteine \u200b\u200bsau interacțiunea lor cu alte proteine \u200b\u200bse schimbă.
Există o altă clasă de receptori cu activitate de proteină kinaza - aceștia sunt receptori conjugați cu proteine \u200b\u200bkinaza. Le lipsește un domeniu catalitic intracelular, dar când interacționează cu un agonist, se leagă sau activează protein kinazele intracelulare pe suprafața interioară a membranei. Aceștia sunt receptori pentru factorii neurotrofici și receptorii care recunosc antigenul pentru limfocitele T și B constând din mai multe subunități. Acestea din urmă interacționează, de asemenea, cu fosfatoza fosfotirozinei. Funcția altor receptori care nu au un domeniu efector intracelular poate fi mediată de unele alte proteine \u200b\u200befectoare.
Alți receptori cu activitate enzimatică proprie au o structură similară. Acestea includ, de exemplu, receptorii cu propria lor activitate de fosfotirozină fosfatază: domeniul lor extracelular este similar în secvența de aminoacizi cu moleculele de adeziune. Pentru mulți receptori cu propria activitate de fosfatazină a fosfotirozinei, liganzii endogeni nu sunt cunoscuți. Cu toate acestea, conform studiilor genetice și biochimice efectuate pe diferite tipuri de celule, activitatea enzimatică a acestor receptori joacă un rol important. Domeniul intracelular al receptorilor hormonali natriuritici atriali, alți NP-uri și, de asemenea, receptorii guanilinei are propria activitate de guanilat ciclază și sintetizează cGMP, care acționează ca un al doilea mediator. Poate există și alți receptori cu propria activitate enzimatică.
Receptorii cuplati cu canal ionic
Receptorii unor mediatori sunt asociați direct cu canalele ionice, atunci când interacționează cu un ligand, trec selectiv anumiți ioni prin membrana celulară (canale chimosensibile, canale receptor ionotropic, receptori ionotropi).
Receptorii cuplati cu proteina G
Aceasta este o clasă destul de mare de receptori care interacționează cu efectorii prin proteine \u200b\u200bG (proteine \u200b\u200bcare utilizează substituția difosfatului de guanină (GDF) pentru trifosfat de guanină (GTP). Acestea includ receptori pentru multe amine biogene, molecule de semnalizare a lipidelor (în special eicosanoide) și diverse peptide și liganzi proteici.Enzimele (adenilat ciclază, fosfolipază C) și canalele membranei de potasiu și calciu acționează ca efectori.Numărul mare și rolul fiziologic important al receptorilor cuplat la proteinele G le face excelente. Obiectivele mele pentru medicamente: aproximativ jumătate din toate medicamentele prescrise de medici (exclusiv antibiotice) acționează asupra acestor receptori.
O celulă poate transporta până la 20 de receptori pe suprafața sa, fiecare dintre ei interacționând selectiv cu unul sau mai multe tipuri de proteine \u200b\u200bG (diferă în diferite tipuri de subunități α). Subunitatea α este capabilă să interacționeze cu una sau mai multe proteine \u200b\u200befectoare, ceea ce vă permite să coordonați semnale de la receptorii diferiților liganzi folosind o proteină G. Pe de altă parte, un singur receptor poate declanșa mai multe mecanisme de transmitere a semnalului intracelular, activând mai multe tipuri de proteine \u200b\u200bG și poate acționa asupra diferitelor proteine \u200b\u200befectoare prin aceeași subunitate α. Un astfel de sistem complex de divergență și convergență a semnalelor oferă o reglare flexibilă a funcțiilor celulare (Ross, 1992).
Receptori intracelulari
Receptorii hormonilor steroizi și tiroidieni, calcitriolului și retinoizilor sunt proteine \u200b\u200bsolubile intracelulare care leagă ADN-ul care reglează transcrierea anumitor gene (Mangelsdorf și colab., 1994). Acești receptori aparțin superfamiliei regulatorilor transcripționali sensibili la ligand. Funcția factorilor de transcripție este reglată de fosforilare, interacțiunea cu proteinele celulare, metaboliții și alte componente reglatoare ale celulei.
A doua sisteme intermediare
cAMP.Sistemele intermediare secundare sunt de asemenea implicate în integrarea semnalelor externe. Deși există mult mai mulți receptori cunoscuți și molecule de semnalizare a proteinelor decât cei de-al doilea mediatori, aceștia din urmă sunt implicați în multe căi din transmisia semnalului celular. Cel mai studiat al doilea intermediar include cAMP, cGMP, Ca 2+, IF 3 (inozitol trifosfat), DAG (diacilglicerol), NO. Acest grup de compuși eterogeni este în continuă creștere. Al doilea mediator interacționează direct (schimbându-și reciproc metabolismul) sau indirect (acționând asupra acelorași ținte intracelulare). Funcția celor de-al doilea mediatori, precum și reglarea formării (sau eliberării) acestora, clivajului și excreției din celulă, sunt luate în considerare în mod convenabil cu exemplu de cAMP. Acest al doilea mediator este sintetizat sub influența cenazelor adenilate la activarea multor receptori conjugați cu proteinele G. Proteina G activează adenilatul ciclază, proteina G i inhibă.
Există cel puțin 10 izoforme de adenilat ciclotază specifice țesuturilor care diferă în mecanismele de reglare a activității.
De regulă, cAMP activează protein kinazele A (proteine \u200b\u200bkinazele dependente de cAMP), un grup mic de proteine \u200b\u200bînrudite. Aceste proteine \u200b\u200bkinaze, la rândul lor, fosforilează nu numai țintele intracelulare finale (enzime, proteine \u200b\u200bde transport), ci și alte proteine \u200b\u200bkinaze și alte proteine \u200b\u200bregulatoare. Acestea din urmă includ, de exemplu, factori de transcripție. Ei sunt responsabili pentru reglarea transcripției genice mediată de cAMP, oferind un răspuns celular întârziat la semnal. Pe lângă activarea proteinelor kinazelor, cAMP acționează direct asupra canalelor de membrană cationică, care joacă un rol important, în special în funcționarea neuronilor. Astfel, semnalul de la cAMP provoacă un lanț de modificări biochimice în celula țintă.
Calciu. Un al doilea mediator bine studiat este Ca 2+ intracelular. Ionii de Ca 2+ intră în citoplasmă în diferite moduri: de-a lungul canalelor de membrană (dependente de proteinele G, dependente de tensiune, reglate de K + sau Ca-Ca 2+), precum și prin canale situate în zone speciale ale reticulului endoplasmic și care se deschid sub acțiunea IF 3, și în mușchiul scheletului ca urmare a depolarizării membranei. Îndepărtarea calciului din plasma citosolică are loc în două moduri: este absorbită de reticulul endoplasmatic sau excretat din celulă. Ca 2+ transmite semnale unui număr mult mai mare de proteine \u200b\u200bdecât cAMP - enzime implicate în metabolismul celular, protein kinazele, proteinele care leagă calciul. Acestea din urmă interacționează cu alți efectori finali și intermediari.
Reglementarea receptorului
Receptorii nu numai că controlează funcțiile fiziologice și biochimice, dar servesc și ca obiecte de reglare. Această reglare se realizează la nivelul sintezei și descompunerii macromoleculelor lor, prin formarea de legături covalente cu alte molecule, interacțiunea cu proteinele regulatoare și mișcarea receptorilor. Transformarea proteinelor și a proteinelor efectoare sunt, de asemenea, supuse reglementărilor. Semnalele de reglementare pot proveni din căile de transmitere intracelulară activate prin stimularea receptorului însuși (prin intermediul unui mecanism de feedback), precum și de la alți receptori (direct sau indirect).
Stimularea pe termen lung a receptorilor medicamentului duce de obicei la o scădere a reacției la acesta - la aceeași concentrație, medicamentul provoacă un efect mai puțin pronunțat. Acest fenomen, numit desensibilizare, refractar, adictiv, joacă un rol important în practica clinică: de exemplu, cu utilizare prelungită Agoniștii β-adrenergici pentru tratamentul pacienților cu AD se reduce severitatea reacției la aceste medicamente.
Desensibilizarea homologică se aplică numai receptorilor stimulați și este specifică pentru ligand. Odată cu desensibilizarea heterologă, scade severitatea reacției la alți liganzi, ai căror receptori acționează prin aceeași cale de transmitere a semnalului intracelular. În primul caz, feedback-ul negativ este furnizat de efectul asupra receptorului în sine (fosforilare, proteoliză, sinteză scăzută), în al doilea caz, pe lângă receptor, poate afecta și alte proteine \u200b\u200bimplicate în transmiterea semnalului intracelular.
Dimpotrivă, dacă receptorii nu sunt stimulați mult timp, sensibilitatea lor la agoniști crește (de exemplu, cu un tratament prelungit cu β-adrenoblocker propronolol, sensibilitatea receptorilor β-adrenergici la β-adrenostimulanți crește).
Tulburări datorate funcției receptorului afectat
Pe lângă diferențele individuale de sensibilitate la medicamente, există boli cauzate de disfuncția anumitor componente ale mecanismului de transmitere a semnalului intracelular de la receptor la efector. Odată cu pierderea funcției receptorilor extrem de specializați, manifestările fenotipice ale bolii pot fi limitate (de exemplu, cu feminizarea testiculelor asociate cu o absență genetică sau cu defecte structurale ale receptorilor androgeni). Dacă este încălcat un mecanism mai universal în interiorul transmiterii semnalului celular, simptomele bolii sunt mai diverse, cum ar fi, de exemplu, cu miastenia gravis și unele forme de diabet zaharat rezistent la insulină, cauzate, respectiv, de disfuncțiile autoimune ale receptorilor N-colinergici și ale receptorilor de insulină. Defectele oricărei componente implicate în transducția semnalului de la mulți receptori duc la tulburări multiple ale endocrinei. Un exemplu este forma heterozigotă a deficitului de proteine \u200b\u200bG care activează adenilatul ciclază în toate celulele (Spiegel și Weinstein, 1995). O formă homozigotă de deficiență a acestei proteine \u200b\u200bpoate duce la deces.
Tulburările din structura sau localizarea receptorilor se pot manifesta ca o reacție slăbită sau îmbunătățită la medicament, precum și alte efecte nedorite.
Mutațiile care codifică receptorii genici sunt capabili să schimbe atât răspunsul la o singură utilizare a medicamentului, cât și eficacitatea tratamentului pe termen lung. De exemplu, un defect al receptorilor β-adrenergici responsabili de relaxarea mușchilor netezi ai bronhiilor și reglarea rezistenței căilor respiratorii agravează scăderea sensibilității acestor receptori la β-adrenostimulanți în timpul tratamentului îndelungat al pacienților cu AD. Pe măsură ce mutațiile responsabile pentru funcția de receptor afectată sunt identificate și genele corespunzătoare sunt clonate, va fi posibil să se dezvolte metode pentru tratarea unor astfel de boli.
Clasificarea receptorilor
În mod tradițional, receptorii medicamentului au fost identificați și clasificați pe baza efectelor și activității relative ale agoniștilor selectivi (stimulanților) și antagoniștilor (blocanților) care acționează asupra acestor receptori. De exemplu, efectele acetilcolinei, care sunt reproduse atunci când interacționează cu receptorii colinergici ai alcaloidului muscarin și sunt blocate de atropină, se numesc efecte muscarinice, iar efectele care sunt reproduse atunci când interacționează cu receptorii colinergici ai nicotinei se numesc efecte nicotinice. Receptorii care mediază efectele muscarinei și nicotinei sunt numiți receptori colinergici M și N, respectiv. Deși o astfel de clasificare nu reflectă de obicei mecanismul de acțiune al medicamentelor, este convenabil pentru sistematizarea efectelor acestora. Într-adevăr, afirmația conform căreia un medicament stimulează receptorii de un anumit tip, în același timp determină spectrul efectelor acestui medicament și substanțe care îmbunătățesc sau slăbesc aceste efecte. Cu toate acestea, validitatea unor astfel de revendicări se poate schimba odată cu identificarea de noi tipuri și subtipuri de receptori, descoperirea unor mecanisme suplimentare de acțiune a medicamentelor sau a efectelor secundare necunoscute anterior.
Subtipuri receptor
Odată cu apariția unei varietăți din ce în ce mai mari de medicamente extrem de selective, a devenit clar că tipurile de receptori cunoscute anterior sunt împărțite în multe subtipuri. Metodele de clonare moleculară au devenit un ajutor semnificativ în studiul noilor subtipuri de receptori, iar prepararea receptorilor recombinanti a facilitat crearea de medicamente care acționează selectiv asupra acestor receptori. Subtipuri diferite, dar înrudite de receptori adesea (deși nu întotdeauna) interacționează cu diferiți agoniști și antagoniști. Receptorii pentru care nu au fost identificați agoniști sau antagoniști selectivi, de obicei nu aparțin unui singur subtip, ci isoformelor aceluiași receptor.Tipurile separate pot diferi, de asemenea, în mecanismele de transmitere a semnalului intracelular. Receptorii colinergici M 1 și M 3, de exemplu, acționează prin proteina G q, care activează fosfolipasa C, determină indirect eliberarea de Ca 2+ din depozitele intracelulare și de receptorii colinergici M 2 și M 4 prin proteina G i, care inhibă adenilatul ciclazei. În același timp, împărțirea receptorilor în tipuri și subtipuri este adesea determinată nu de mecanismul de acțiune, ci de o alegere aleatorie sau se bazează pe idei consacrate. Deci, receptorii α 1 -, α 2 - și β-adrenergici diferă ca răspuns la medicamente și în transmiterea semnalului (activează proteinele G i, G q și G s), deși receptorii α și β-adrenergici sunt de diferite tipuri, și α 1 - și α 2 -adrenoreceptori - la subtipuri diferite din același tip. Izoformele α -adrenoreceptorilor α 1A, α 1B și α 1D diferă puțin în proprietățile lor biochimice; același lucru este caracteristic pentru subtipurile de izoformrasă ale receptorilor β-adrenergici (β 1, β 2 și β 3).
Diferențele dintre subtipurile de receptori sunt utilizate pentru a crea medicamente extrem de selective, de exemplu, medicamente care au efecte diferite asupra aceluiași țesut datorită legării cu subtipuri de receptori care diferă în mecanismele de transmitere a semnalului intracelular. În plus, medicamentele pot viza în mod selectiv anumite celule sau țesuturi care exprimă receptorii unui subtip. Cu cât este mai mare selectivitatea medicamentelor (în raport cu un anumit țesut sau în raport cu un anumit efect), cu atât este mai favorabil raportul dintre beneficiile și efectele nedorite ale acestuia.
Folosind metode genetice moleculare, nu numai izoforme diferite de receptori au fost descoperite, dar și gene care codifică receptori noi, necunoscuți anterior. Mulți dintre acești receptori sunt deja alocați unei clase sau altei clase cunoscute și funcția lor a fost studiată folosind liganzii corespunzători. Cu toate acestea, liganzi nu au fost încă găsiți pentru unii receptori.
Descoperirea multor izoforme ale aceluiași receptor codificate de gene diferite (mai ales dacă izoformele nu diferă în mecanismele de transmitere a semnalului intracelular și interacționează cu aceiași liganzi endogeni) permite ca expresia receptorilor din celule diferite să fie reglementată independent în conformitate cu nevoile organismului în diferite perioade de vârstă.
Acțiunea medicamentului mediat non-receptor
Nu toate medicamentele acționează prin structurile macromoleculare - receptori. Unele medicamente interacționează cu molecule mici sau ioni care sunt prezente în organism în mod normal sau într-una sau alta stare patologică. Deci, antiacidele neutralizează acidul clorhidric în stomac. Mesna (un medicament care este excretat rapid de rinichi și neutralizează radicalii liberi) se leagă de metaboliții activi ai unor medicamente anticancerigene, reducând severitatea reacțiilor adverse din tractul urinar. O serie de substanțe biologice inactive (de exemplu manitolul) pot fi introduse în cantități suficiente pentru a crește osmolaritatea fluidelor biologice și, astfel, a schimba distribuția fluidelor extracelulare și intracelulare. Cu ajutorul acestor substanțe este posibilă creșterea diurezei, creșterea bccului, eliminarea edemului cerebral. În plus, sunt utilizate ca laxative.
Unele medicamente se pot integra în componentele celulei și își pot schimba funcțiile datorită asemănărilor structurale cu substanțele care compun aceste componente. De exemplu, analogii purinelor și pirimidinelor sunt inserate în acizi nucleici și sunt folosiți ca agenți antivirali și antitumori.
AP Viktorov "Farmacologie clinică"
De regulă, mecanismul de acțiune al medicamentelor se bazează pe capacitatea lor de a iniția (declanșa) procese biochimice complexe n / sau biofizice care în final modifică și / sau optimizează activitatea funcțională a celulei țintă.
Medicamentele își pot duce acțiunea împotriva organelor și / sau a celulelor țintă prin:
Interacțiune chimică directă;
Interacțiunea fizico-chimică pe membrana celulară;
Acțiuni asupra enzimelor de specialitate;
Acțiuni privind genele de reglementare;
Acțiuni asupra receptorilor specifici.
Interacție chimică directă LS. Acest mecanism de acțiune al medicamentelor este destul de rar și poate fi realizat în afara celulei, de exemplu, în lumenul stomacului sau al intestinelor. Esența sa constă în faptul că medicamentele intră într-o reacție chimică directă cu molecule și / sau ioni care se formează în organism într-o stare normală când apare o afecțiune patologică. Un exemplu de interacțiune chimică directă este reacția chimică de neutralizare a acidului clorhidric al stomacului când luați medicamente antiacide (vezi T. 2, p. 112).
Interacțiunea fizico-chimică a medicamentelor pe membrana celulară. Una dintre principalele funcții ale membranei citoplasmatice este implementarea schimbului de ioni între citoplasmă și mediul extracelular. Schimbul de ioni transmembranari poate avea loc, de asemenea, prin canale ionice transmembrane speciale dependente de tensiune - sodiu, potasiu, calciu, clor etc. Unele medicamente, care ajung la membrana celulară, interacționează cu aceste canale și le modifică activitatea funcțională. Deci, de exemplu, efectul antiaritmic al unui medicament de clasa IA, chinidina, se bazează pe capacitatea sa de a bloca trecerea ionilor de Na + prin canalele de sodiu transmembranare (vezi T. 2, p. 35).
Efectul medicamentelor asupra enzimelor specializate. O cantitate relativ mică de medicamente își realizează efectul farmacologic prin schimbarea activității unor enzime celulare specializate. Medicamentele care cresc activitatea enzimelor celulare se numesc inductori de enzime. O astfel de acțiune este deținută, de exemplu, de somnifere și de medicamentul anticonvulsivant fenobarbital, care îmbunătățește semnificativ activitatea enzimelor hepatice microsomale. Semnificația biologică a acestui efect al fenobarbitalului și LS aproape de acesta va fi luată în considerare mai jos.
Medicamentele care inhibă activitatea enzimelor specializate se numesc inhibitori ai enzimei. Deci, de exemplu, un antidepresiv din grupul inhibitorilor monoaminoxidazei (MAO), medicamentul pirlindol își realizează efectul antidepresiv prin suprimarea activității enzimei MAO în sistemul nervos central (vezi T. 1, p. 294).
Capacitatea de a inhiba activitatea enzimei acetilcolinesterază stă la baza activității farmacologice a medicamentelor anticolinesterază, de exemplu fizostigmină. Se știe că în condiții fiziologice, acetilcolinesteraza inactivează (distruge) acetilcolina, un neurotransmițător care transmite excitația în sinapsele sistemului nervos parasimpatic. Physostigmine, care suprimă activitatea acetilcolinesterazei, promovează acumularea în sinapsele sistemului parasimpatic al neurotransmițătorului acetilcolină, ca urmare a creșterii tonusului sistemului nervos parasimpatic, care se manifestă la nivel sistemic prin dezvoltarea bradicardiei, scăderea tensiunii arteriale (BP), a gastro și cardiacă elev etc.
Medicamentele pot interacționa reversibil și ireversibil cu enzimele. De exemplu, medicamentul enalapril inhibă reversibil activitatea enzimei de conversie a angiotensinei, ceea ce implică, în special, o scădere a tensiunii arteriale, în timp ce substanțele toxice organofosforice inhibă ireversibil activitatea acetilcolinesterazei.
Efectul medicamentelor asupra genelor de reglementare. În prezent, oamenii de știință fac încercări de a crea medicamente care să realizeze efectele lor farmacologice, afectând direct activitatea fiziologică a genelor reglatoare. Această tendință pare deosebit de promițătoare după ce structura genomului uman a fost descifrată în 2000. Se crede că normalizarea selectivă a funcției genelor reglatoare sub influența medicamentelor va face posibilă obținerea unui succes în tratamentul multor boli, inclusiv anterior incurabile.
Efectul medicamentelor asupra receptorilor. Înainte de a trece la specificul interacțiunii medicamentelor cu receptorii, este necesar să clarificăm ce înțelegem prin termenul „receptor” (din latinescul receptio - ia, ia).
De la cursul fiziologiei se știe că termenul „receptor” înseamnă formațiuni foarte specializate care sunt capabile să perceapă, să transforme și să transmită energia unui semnal extern către sistemul nervos. Astfel de receptori sunt numiți senzorial (din lat. Sensus - senzație, senzație, percepție).
Receptorii senzoriali includ receptorii organelor auzului, vederii, mirosului, gustului, atingerii etc. Receptorii senzoriali ai acestor organe aparțin așa-numitilor exteroreceptori.
Dacă prezența organelor senzoriale care răspund la stimuli externi de iritație este cunoscută încă din cele mai vechi timpuri, atunci prezența receptorilor senzoriali în interiorul corpului a fost pusă la îndoială până la mijlocul secolului al XIX-lea. Pentru prima dată, prezența unor astfel de receptori în interiorul corpului a fost sugerată de fiziologul rus I.F.Pion, care a arătat o scădere a tensiunii arteriale în 1866 din cauza iritării aortice într-un experiment cu iepure. Această descoperire a dat naștere căutării și studiului receptorilor localizați în interiorul corpului, iar acești receptori înșiși au fost numiți interoreceptori.
Până la începutul secolului XX a fost dezvăluit un număr suficient de interoreceptori senzoriali și s-a dovedit rolul lor important în reglarea funcțiilor fiziologice ale organismului.
În 1905, J. Langley a dovedit că atunci când un medicament este aplicat pe o membrană celulară, un efect farmacologic se dezvoltă dacă este aplicat doar pe o anumită zonă a acestuia. Mai mult, acest site nu constituie decât o mică parte din suprafața totală a suprafeței celulare. Această observație a permis lui J. Langley să concluzioneze că pe membrana celulară există site-uri de receptori specializați care interacționează cu medicamente.
Cu toate acestea, prioritatea în crearea teoriei receptorilor a acțiunii medicamentelor aparține fiziologului german P. Ehrlich, care în 1906 a introdus pentru prima dată termenul de „receptor” și a formulat postulatul „medicamentul nu funcționează dacă nu este fixat pe membrana celulară”. Conform teoriei lui P. Ehrlich, o moleculă de medicament are două grupe active din punct de vedere funcțional, dintre care una asigură fixarea acesteia pe suprafața celulelor din regiunea receptorului de droguri, iar cea de-a doua grupă funcțională interacționează cu receptorul și declanșează un lanț complex de reacții biochimice care îi modifică activitatea fiziologică (celulară). .
Astfel, încă de la începutul secolului XX. a devenit evident că există cel puțin două clase de interoreceptori: receptorii senzoriali care transmit informații despre starea organelor interne și a țesuturilor corpului către sistemul nervos central; etichetarea receptorilor care interacționează cu medicamentele care modifică activitatea funcțională a celulelor țintă.
Trebuie menționat imediat că, în viitor, în textul manualului, pentru a evita confuziile în terminologie, receptorii pentru medicamente și substanțele biologic active, adică. etichetat sau citoreceptori. va fi notat de termenul „receptor”, în timp ce interoreceptorii senzoriali vor fi notați de un termen care caracterizează activitatea lor funcțională, de exemplu, „baroreceptori”, „receptori ai durerii” etc.
Descoperirea de către P. Ehrlich pe membrana celulară a receptorilor medicamentelor a servit ca punct de plecare pentru dezvoltarea științei farmacologice, în special farmacodinamica, una dintre principalele sarcini ale căreia este studierea mecanismelor receptorilor de acțiune a medicamentelor.
În prezent, a fost dezvăluită structura unui număr mare de receptori celulari, caracteristicile interacțiunii anumitor compuși biologic activi cu aceștia, ceea ce a făcut posibilă, pe de o parte, înțelegerea mecanismului de acțiune a medicamentelor cunoscute, iar pe de altă parte, a fost baza pentru crearea de noi medicamente extrem de eficiente.
În mod natural, este greu de imaginat că, în cursul evoluției, în corpul uman s-au format receptori pentru diverse medicamente sintetice (obținute chimic), mai ales că marea majoritate a medicamentelor prezentate pe piața farmaceutică modernă au fost sintetizate în ultimii 50 de ani sau mai puțin. Este dovedit că aparatul receptor al celulei este o formație funcțional-structurală foarte veche. Deci, a- și β-adrenergici (receptori, a căror interacțiune norepinefrină și adrenalină afectează activitatea funcțională a celulei) se găsesc nu numai în celulele animale, ci și pe membranele celulare ale celulelor plantelor, de exemplu, în celulele plantei nittella, unde a- și β- adrenorecentrele reglează mișcarea protoplasmei (conținutul celular).
Atunci care sunt receptorii pentru medicamentele descoperite de P. Ehrlich și de ce interacționează cu aceștia?
În prezent, nu există nici o îndoială că așa-numiții receptori de droguri sunt de fapt receptori pentru substanțele biologice active endogene (produse în organism) implicate în reglarea activității funcționale a organelor interne și a țesuturilor corpului. Astfel de compuși activi biologic includ substanțe eliberate de terminațiile nervoase la momentul transmiterii semnalului nervos, precum și hormoni, vitamine, aminoacizi etc. Pentru fiecare substanță biologică activă endogenă, există receptori strict specifici pentru aceasta. Deci, de exemplu, substanța biologică activă produsă în organism, adrenalina, poate activa strict a- și β-adrenoreceptorii, iar glucocorticosteroizii - hormoni ai cortexului suprarenal - interacționează numai cu receptorii glucocorticosteroizi strict specific pentru ei.
Medicamentele sintetice care își realizează efectele prin interacțiunea cu aparatul receptor al celulei, în structura lor chimică, sunt mai mult sau mai puțin similare cu compușii activi biologic endogeni care interacționează cu receptorii similari. De exemplu, medicamentele sintetice cu vasoconstrictor (care provoacă vasoconstricție), fenilefrina este apropiată în structura sa chimică de substanța endogenă biologic activă norepinefrină, prin urmare, la fel ca norepinefrina, are capacitatea de a stimula a-adrenoreceptorii.
Uneori, datorită particularităților structurii lor chimice, medicamentele pot interacționa nu cu receptorul în sine, ci cu porțiunea adiacentă a membranei celulare. Întrucât, în acest caz, medicamentul nu interacționează cu receptorul în sine, ci cu porțiunea adiacentă a membranei celulare, acestea nu vorbesc despre un efect excitant sau de blocare asupra receptorului, ci despre un efect sau efect alosteric (din grecescul alios - alt, diferit). Ca urmare, poate apărea o modificare atât a structurii membranei adiacente receptorului, cât și a componentelor individuale ale receptorului în sine, ceea ce poate atrage o schimbare a sensibilității receptorului față de o substanță biologică activă specifică acestuia. În cazurile în care sensibilitatea receptorului la o substanță activă biologic crește, vorbesc de sensibilizare (din latinescul sensus - sens) sau sensibilizare (din latinescul sensibilis - sensibilitate) al receptorului și în cazurile în care sensibilitatea receptorului scade, vorbesc despre desensibilizare. receptor.
Particularitatea efectului alosteric constă în faptul că medicamentele care au acest tip de mecanism de acțiune nu afectează în mod direct transmiterea unui impuls nervos, ci îl modifică în direcția dorită. De exemplu, mecanismul de acțiune al anxioliticelor (medicamente anti-anxietate; sinonim: tranchilizanți), care în structura lor chimică sunt derivați ai benzodiazepinei, se bazează pe fenomenul excitării alosterice a receptorilor postsinaptici de benzodiazepină. Excitația acestuia din urmă, la rândul său, promovează activarea receptorilor inhibitori postsinaptici ai acidului gamma-aminobutiric (receptorii GABA), care se manifestă clinic prin eliminarea simptomelor bolilor nevrotice precum anxietatea, anxietatea, frica etc.
Receptorii, care interacționează cu care o substanță sau un medicament biologic activ schimbă în orice fel starea funcțională a unei celule țintă, sunt numiți specifici.
În plus față de receptorii specifici, așa-numiții receptori specifici medicamentului sunt izolați. În literatura medicală de specialitate, acești receptori mai sunt numiți „locul pierderii” de medicamente. Prin contactarea unor astfel de receptori, medicamentele nu au niciun efect biologic, dar ele însele devin biologice inactive. Un exemplu de acest tip de receptori poate servi ca receptori localizați pe proteine \u200b\u200bplasmatice, în special, pe proteine \u200b\u200bsolubile în apă - albumina. Semnificația acestui fenomen va fi discutată în detaliu mai jos (vezi T. 1, p. 72).
Structura receptorilor este destul de complexă, dar majoritatea sunt macromolecule proteice sau glicoproteine, care pot include și ioni, lipide, acizi nucleici etc. Receptor i.e. macromolecula de proteine \u200b\u200bcare o formează se caracterizează printr-o aranjare spațială specifică pentru fiecare receptor, a grupelor sale chimice. Macromolecula proteică care formează receptorul poate fi integrată (cufundată) în stratul lipidic al membranei citoplasmatice sau localizată în interiorul celulei. Principala funcție a unui receptor de celule este „recunoașterea” unui semnal chimic transmis acestuia printr-o substanță endogenă biologic activă și / sau medicamente și transformarea acestuia în răspunsul biochimic și / sau biofizic corespunzător al celulei.
Se credea anterior că medicamentele sau substanțele endogene active biologic interacționează cu receptorii de tip „cheie și blocare”, adică. receptorul are o astfel de structură care permite medicamentului să-ți găsească receptorul „tău”, să se conecteze la acesta și, așa cum era, „pornește” și „oprește-l”. Cu toate acestea, odată cu dezvoltarea științei medicale, a devenit evident că ego-ul nu este chiar așa. În prezent, procesele moleculare de conversie a semnalelor extracelulare în celule intracelulare, care reglează funcția celulelor, au fost deja studiate destul de bine. mecanisme care au ca efect efectul interacțiunii substanțelor endogene active sau a medicamentelor cu receptorii.
Atunci când interacționează cu receptorul unei substanțe endogene biologic active și / sau o L C activă, apare o conformație - o schimbare spațială sub forma unei macromolecule proteice, care este declanșatorul pentru diverse procese intracelulare care determină răspunsul unei celule țintă la un mediator și / sau un medicament. De exemplu, activarea receptorilor adrenergici bronșici netede sub influența fenotolului β2-adrenostimulator conduce la o creștere a activității enzimei adenilate ciclază, ceea ce contribuie la acumularea de adenozin monofosfat ciclic (cAMP) în celulă și, ca urmare, relaxarea celulară.
În termeni biologici generali, receptorii celulari pot fi considerați ca „organe senzoriale” strict specializate ale celulelor, prin intermediul cărora percep „informații” care emană, de exemplu, din sistemul nervos central și / sau din sistemul endocrin. În ciuda rolului important al aparatului receptor, receptorii ocupă doar o parte nesemnificativă a membranei celulare. De exemplu, aparatul receptor M-colinergic al unei celule nu ocupă mai mult de 1/6 000 din suprafața sa.
Studierea caracteristicilor interacțiunii medicamentelor cu receptorul, pe de o parte, ne permite să înțelegem baza mecanismului molecular al acțiunii sale, iar pe de altă parte, oferă informații despre ce schimbări ar trebui făcute în structura medicamentelor pentru a spori capacitatea acestuia de a interacționa cu acest receptor, adică. . permite sinteza țintită de noi medicamente extrem de eficiente.
În condiții fiziologice, diferiți receptori celulari nu funcționează independent, ci sunt într-o interacțiune constantă între ei, reglând astfel activitatea specifică a celulei. De exemplu, activarea β-adrenoreceptorilor cardiaci prin norepinefrină endogenă determină, în special, o creștere a numărului de contracții cardiace și activarea receptorilor M-colinergici cardiaci de acetilcolina endogenă, dimpotrivă, determină o scădere a numărului de contracții cardiace.
O mare contribuție la înțelegerea mecanismelor receptorilor de acțiune a medicamentelor a fost realizată prin descoperirea receptorilor pre și post-sinaptici. Sinapsa (din greacă sinapsis - conexiune, conexiune) este o zonă de contact specializată între celulele nervoase sau alte structuri excitabile ale corpului, care asigură transmiterea informațiilor primite și păstrarea semnificației sale informaționale. Studiul structurii și rolului funcțional al sinapselor a fost început la sfârșitul secolului XIX. după aceea, istologul spaniol S. Ramon n Cajal (S. Ramon la Cajal) a sugerat prezența unui sistem de transmisie specializat în sistemul nervos central. Sinapsele și-au luat numele în 1897, când fiziologul englez C. Sherrington a propus acest termen să se refere la zona de contact dintre celulele nervoase.
În prezent, există trei tipuri de sinapse:
1) sinapse „electrice” în care se transmit informații prin transferul unui semnal electric dintr-o membrană pre-sinaptică. Acest tip de sinapsă se numește efaps (din greacă. Eppsis - contact strâns);
2) sinapse „chimice” în care informațiile sunt transmise prin substanțe biologice speciale active - neurotransmițători (din greacă. Neuron - nervos și latin. Mediator - mediator; sinonim: mediator);
3) sinapsele „mixte” în care informațiile sunt transmise atât chimic cât și electric.
Efectele farmacologice din marea majoritate a medicamentelor care afectează funcțiile sinapselor sunt realizate prin efectul lor asupra gothului sau a unei alte etape de transmitere a semnalului în sinapsele chimice, adică. în sinapsele de al doilea fel.
De regulă, sinapsele chimice sunt clasificate de neurotransmițătorii care transmit impulsuri nervoase în ele, astfel:
Sinapsele în care acetilcolina acționează ca mediator sunt numite colinergice;
Sinapsele în care adrenalina și norepinefrina acționează ca mediator sunt numite adrenergice;
Sinapsele în care ATP și adenozina acționează ca mediator sunt numite purinergice;
Sinapsele în care acidul gamma-aminobutiric acționează ca mediator se numesc GABA-ergic etc.
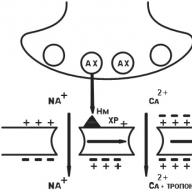
În prezent, structura sinapselor este bine înțeleasă. Sinapsa constă dintr-un proces presinaptic al unei celule nervoase (capătul axonului) și un aparat de recepție „semnal” situat pe membrana unei celule efectoare („executive”).
Axonul neuronului eferent, care se apropie de celula efectoare, pierde teaca de mielină, se extinde și formează așa-numitul îngroșare presinaptică (Fig. 1.5). Suprafața nervului care se termină cu fața membranei celulare a celulei efectoare se numește membrană presinaptică. Locul celulei efectoare opusă membranei presinaptice se numește membrană postsinaptică (vezi Fig. 1.5). În funcție de caracteristicile structurale ale sinapsei, membrana presinaptică poate avea mai multe sau mai puține pliuri și are astfel o suprafață mai mare sau mai mică. În sinapsele chimice, membrana presinaptică nu ia contact direct cu membrana postsinaptică, ci este separată de ea printr-o distanță mică numită fanta sinaptică (vezi Fig. 1.5).
Îngroșarea presinaptică, adică porțiunea finală a axonului conține un număr mai mare de mitocondrii, organele intracelulare implicate în sinteza și acumularea de energie, care este mai mare decât corpul neuronului, ceea ce indică intensitatea proceselor energetice care apar în această secțiune a celulei nervoase. Pe lângă mitocondrii, îngroșările presinaptice conțin un număr mare de vezicule mici - vezicule. În medie, aproximativ 20.000 vezicule sunt conținute într-o îngroșare presinaptică. Acestea din urmă sunt situate neuniform în îngroșarea presinaptică, de regulă, majoritatea sunt situate în apropierea membranei presinaptice. Neurotransmițătorul este sintetizat în corpul și axonul neuronului și se acumulează în vezicule. Fiecare vezicule conține câteva mii de molecule ale unui neurotransmițător (de la I 000 la 50 000). Când apare un impuls nervos, vezicula fuzionează cu membrana presinaptică și neurotransmițătorul este secretat în fanta sinaptică (vezi Fig. 1.5).
Fig. 1.5. Diagrama schematică a structurii sinapsei „chimice”:
a este o imagine schematică; b - micrografie electronică; 1- terminarea nervului presinaptic; 2 - membrană prsinaptică; 3 - membrană postsinaptică; 4 - fanta sinaptică; B - vezicule; NM - neurotransmițător; P - receptor postsinaptic: OZ - captarea „inversă” a unui neurotransmițător; SF este o enzimă specializată care distruge excesul de neurotransmițător în fanta sinaptică
Formațiile receptorilor active sunt localizate pe membrana postsinaptică, care sunt capabile să interacționeze cu neurotransmițătorul eliberat de membrana presinaptică în timpul trecerii unui impuls nervos. Receptorii localizați pe membrana postsinaptică sunt numiți receptori sinaptici sau postsinaptici în literatura medicală de specialitate. Prin „receptori postsinaptici” se înțelege macromoleculele de natură proteică încorporate în membrana postsinaptică cu structură și funcție predeterminate genetic, capabile să interacționeze reversibil cu neurotransmițătorii și / sau medicamente datorită grupurilor funcționale ale centrului activ (partea „recunoscătoare” a macromoleculei).
Transmiterea semnalului nervos în sinapsă are loc după cum urmează: sub influența unui stimul nervos, veziculele se deplasează către membrana presinaptică, iar neurotransmițătorul este secretat de exocitoza în fanta sinaptică (vezi figura 1.5). Neurotransmițătorul eliberat în fanta sinaptică ajunge la membrana postsinaptică, unde, interacționând cu receptorul postsinaptic, declanșează un lanț de reacții biochimice și / sau biofizice, rezultatul căruia este răspunsul fiziologic al celulei țintă. Cu toate acestea, nu toată cantitatea de neurotransmițător eliberat ajunge la receptorii postsinaptici și interacționează cu aceștia. O parte din neurotransmițător este capturată de membrana presinaptică și se „întoarce” la locurile de stocare. Acest fenomen se numește fenomenul de recaptare a neurotransmițătorilor.
Cantitatea rămasă de receptor neurotransmițător care nu interacționează este distrusă în fanta sinaptică de către enzime specializate. Acest fenomen se numește degradarea neurotransmițătorilor. De exemplu, enzima acetilcolinesteraza catalizează (accelerează) procesul de degradare (distrugere) în fanta sinaptică a neurotransmițătorului acetilcolină.
Spre deosebire de neurotransmițător, produsele sale metabolice au activitate neurotransmițător. Întregul proces de interacțiune a neurotransmițătorului cu receptorii și distrugerea excesului său de către o enzimă specifică este extrem de scurt și nu depășește 2 ms (1 ms \u003d 0,001 s).
O astfel de durată scurtă a acestui proces este explicată, pe de o parte, prin eliberarea extrem de rapidă a neurotransmițătorului de la receptor și, pe de altă parte, prin rata mare de inactivare enzimatică a neurotransmițătorului în casca sinaptică.
Activitatea fundamentală funcțională a sinapsei poate fi modificată după cum urmează:
Pentru a accelera, reduce sau bloca sinteza, acumularea și / sau catabolismul (distrugerea) neurotransmițătorului în finalul presinaptic. Drept urmare, conținutul neurotransmițătorului și, în consecință, intensitatea activității sale fiziologice se va schimba cumva.
De exemplu, reserpina simpatolitică previne acumularea de catecolamine în veziculele sinaptice până la golirea lor completă. Ca urmare, cantitatea de neurotransmițător norepinefrină eliberată în fanta sinaptică scade brusc. La nivelul sistemului, acest efect se realizează sub forma scăderii tensiunii arteriale. Unele medicamente nu afectează în mod direct conținutul neurotransmițătorilor în finalul presinaptic, ci inhibă activitatea enzimelor care le distrug. Astfel, o serie de antidepresive acționează. De exemplu, antidepresivul pirlindol inhibă (suprimă) activitatea enzimei monoaminoxidază la capătul presinaptic și, ca urmare, crește concentrația de neurotransmițători precum norepinefrină, dopamină și serotonină în ea. Clinic, acest efect al pirlindolului se manifestă printr-o scădere a sentimentelor de anxietate și frică, îmbunătățirea stării de spirit, creșterea activității fizice, etc;
Schimbă (facilitează, complică) capacitatea neurotransmițătorului de a pătrunde în membrana presinaptică și, prin urmare, crește sau scade cantitatea de neurotransmițător eliberat în fanta sinaptică cu fiecare puls.
De exemplu, amfetamina psiostimulantă facilitează eliberarea de catecolamine în sinapsele adrenergice ale sistemului nervos central și, prin urmare, crește conținutul lor în fanta sinaptică. Clinic, acest efect al medicamentului se manifestă într-o stare de spirit îmbunătățită, o senzație de creștere a forței, performanță crescută. Toxina toxică blochează eliberarea de neurotransmițători inhibitori (GABA, glicină) în sistemul nervos central și, prin urmare, reduce brusc conținutul acestora în fanta sinaptică, care se manifestă clinic prin dezvoltarea de crize;
Blochează sau stimulează recaptarea neurotransmițătorilor de către membrana presinaptică și, prin urmare, crește sau scade concentrația de neurotransmițători în fanta sinaptică.
De exemplu, imipramina antidepresivă triciclică blochează recaptarea neurotransmițătorului norepinefrină de membrana presinaptică și, prin urmare, crește brusc concentrația sa în fanta sinaptică. Clinic, acest efect al imipraminei se manifestă prin starea de spirit îmbunătățită, creșterea activității mentale și fizice;
Stimulează sau blochează activitatea enzimelor care distrug neurotransmițătorul în fanta sinaptică.
De exemplu, fizostigmina medicamentului anticolinesterază reduce activitatea enzimei acetilcolinesterază, care distruge neurotransmițătorul acetilcolină în fanta sinaptică și, prin aceasta, contribuie la o creștere a concentrației sale, care poate fi manifestată clinic, în special, printr-o scădere a presiunii și constricției intraoculare a pupilei.
Stimulează sau blochează receptorii postsinaptici, adică imită sau blochează efectul neurotransmițătorilor.
De exemplu, analgezicele narcotice care excită receptorii opioizi postsinaptici și prin aceasta imită afectarea neurotransmițătorilor - encefaline. Stricnina, prin blocarea receptorilor neurotransmițătorului inhibitor glicină, împiedică realizarea efectului său inhibitor, ca urmare, stricnina în doze mari provoacă convulsii.

Fig. 1.6. Reprezentarea schematică a localizării receptorilor pre și post sinaptici, astfel cum este exemplificată de sinapsa adrenergică (explicație în
NM - neurotransmițător; M2 (-) - heteroreceptor presinaptic colinergic "inhibitor"; β 1 (+) - autoreceptor presinaptic „activator” adrenergic; β - receptor postsinaptic adrenergic
Pe lângă receptorii localizați pe membrana postsinaptică, adică receptori postsinaptici, receptori localizați pe membrana presinaptică, adică receptorii presinatici (Fig. 1.6). În ciuda faptului că atât receptorii pre și post sinaptici pot fi excitați de același neurotransmițător, rolul funcțional al acestor formațiuni de receptori în sinapsele este diferit. Dacă receptorii postsinaptici sunt veriga finală pentru transmiterea unui impuls nervos către un organ efector, adică. asigură conducerea unidirecțională a unui impuls nervos de la centru la periferie, apoi receptorii presinaptici participă la
reglarea activității neurotransmițătorului sinapsei, adică într-o oarecare măsură afectează procesele de eliberare și / sau sinteza unui neurotransmițător în el. Trebuie subliniat faptul că receptorii presinaptici nu participă direct la conducerea unui impuls nervos de la un neuron la un organ efector.
Receptorii presinaptici sunt împărțiți în două grupe mari: receptorii auto- și heteroneuromodulatori (vezi Fig. 1.6).
Autoreceptorii presinaptici includ receptori care sunt excitați de propriul neurotransmițător pentru această sinapsă.
De exemplu, în sinapsele localizate în zona de contact dintre nervii somatici și mușchiul striat, când neurotransmițătorul conține un exces de acetilcolină în fanta sinaptică, interacționând cu autoreceptorii presinaptici, inhibă eliberarea unei noi porțiuni de neurotransmițător. excitarea autoreceptorilor presinaptici reglează eliberarea acetilcolinei din terminalele presinaptice.
Cu toate acestea, pe membrana presinaptică, pe lângă autoreceptori, adică. receptorii care sunt sensibili la un neurotransmițător care transmite excitația într-o sinapsă dată, pot fi localizați receptori care nu sunt sensibili la un neurotransmițător care transmite excitația într-o sinapsă dată, dar interacționează cu un alt tip de neurotransmițător.
De exemplu, pe membrana presinaptică sinaptică, în care acetilcolina este neurotransmițătorul, se pot localiza receptori presinaptici sensibili la neurotransmițătorul norepinefrină. Acest tip de receptor presinaptic se numește receptor heteroneuromodulant.
Astfel, sinapsa este o formațiune anatomică și funcțională complexă care asigură transmiterea unui impuls nervos de la un neuron la un neuron sau de la un neuron la o celulă efectoare.
Secvența activității funcționale a sinapsei (etapele transmiterii sinaptice) este următoarea:
Sinteza și acumularea unui neurotransmițător în vezicule localizate în îngroșări presinaptice (sinteza unui neurotransmițător apare nu numai la îngroșările presinaptice, ci și la un neuron și axoni);
Eliberarea neurotransmițătorului în fanta sinaptică în momentul trecerii unui impuls nervos;
Interacțiunea neurotransmițătorului cu receptorii postsinaptici, care implică activarea receptorilor și o modificare a activității funcționale a celulei efectoare;
Inactivarea neurotransmițătorului (enzimatic) și / sau recaptării acestuia de către membrana presinaptică, adică. refacerea capacității sinapsei de a transmite din nou un impuls nervos unei celule efectoare.
Sinapsele au următoarele proprietăți de bază:
Conducerea unilaterală a excitației (un impuls nervos poate trece doar de la membrana presinaptică la cea postsinaptică);
Întârziere sinaptică, adică un anumit timp este alocat pentru transmiterea unui impuls nervos în sinapsă. (Viteza de transmitere sinaptică este, în medie, de peste 10 ori mai mică decât viteza de propagare a unui impuls nervos prin nerv. Pentru o sinapsă chimică, aceasta de obicei variază între 0,2-0,5 ms);
Oboseală - o scădere treptată sau încetarea completă a transmiterii unui impuls nervos cu stimulare nervoasă prelungită. Baza acestui fenomen este, pe de o parte, epuizarea rezervelor de neurotransmițător în îngroșări presinaptice, iar pe de altă parte, o scădere a sensibilității la receptorii postsinaptici la neurotransmițător;
Sensibilitate ridicată a formațiunilor sinaptice la medicamente și otrăvuri.
Pe ultima proprietate a sinapselor se bazează întreaga farmacologie a medicamentelor care influențează activitatea funcțională a sinapselor localizate în diferite organe și țesuturi ale corpului. Trebuie subliniat faptul că obiectul acțiunii farmacologice poate fi oricare dintre etapele de transmitere sinaptică. Ca medicamente care afectează transmisia sinaptică, sunt folosiți analogi exogeni ai neurotransmițătorilor, precursorilor lor chimici și a altor substanțe biologic active care pot modifica în orice mod activitatea funcțională a sinapsei.
Trebuie menționat că multe medicamente nu au unul, ci mai multe puncte de aplicare a efectului la nivelul sinapselor. Deci, de exemplu, pirlindolul antidepresiv nu numai că inhibă activitatea enzimei monoaminoxidazei în fanta sinaptică, ci și blochează recapitularea norepinefrinei de către membrana presinaptică.
În legătură cu localizarea receptorului în sinapsă, aceștia pot fi împărțiți în presinaptici, postsinaptici și extrasinaptici. Acestea din urmă, de exemplu, includ receptori localizați pe membranele celulare ale trombocitelor.
Din punct de vedere al topografiei celulare (locație), receptorii pot fi, de asemenea, clasificați în funcție de locația lor pe structurile celulare, după cum urmează:
receptori ai membranei - receptori localizați pe membrana citoplasmatică;
receptori citosolici - receptori localizați pe formațiuni intracelulare;
receptori nucleari - receptori localizați pe membrana nucleului celular.
După cum sa menționat anterior, ca urmare a interacțiunii cu receptorul substanțelor sau medicamentelor biologice endogene, activitatea funcțională a celulelor țintă se modifică. Acest proces poate fi implementat în moduri diferite, strict definite pentru diferite tipuri de receptori. În conformitate cu aceasta, în prezent se disting patru tipuri de receptori, fiecare având propriul său mecanism, diferit fundamental de celelalte, prin care un semnal de la receptor inițiază o cascadă de reacții biochimice și / sau biofizice care conduc la o schimbare a stării funcționale a celulelor țintă.
Primele trei tipuri de receptori sunt localizați pe membrana celulară (citoplasmică), iar al patrulea tip de receptori include receptorii citosolici și nucleari.
Receptorii de tip I includ receptorii celulari (cu membrană) care își realizează efectele prin așa-numitele proteine \u200b\u200bG de semnalizare (Fig. 1.7).
În prima etapă, o substanță sau medicament biologic activ, „urcând” către membrana celulară, „recunoaște” receptorul și interacționează cu acesta, după care receptorul activează un semnal specializat proteină G, localizat pe suprafața interioară a membranei. Mai mult, proteina G activată modifică activitatea funcțională a elementului efector intern, care, de regulă, este enzimele. Apoi elementul efector, care este o enzimă, activează un mesager secundar sau un mesager secundar, care declanșează o cascadă de reacții biochimice care schimbă activitatea funcțională a celulelor țintă.
Receptorii de celule de tip I, adică receptorii conjugați cu proteine \u200b\u200bG semnal sunt similare structural unul cu celălalt, iar în organizarea lor spațială sunt structură serpentină (din franceză serpantină - șarpe, bilă) (Fig. 1.8).
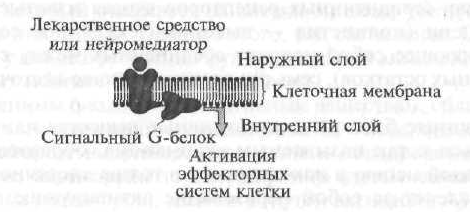
Fig. 1.7. Structura receptorului de tip 1 (explicație în text)
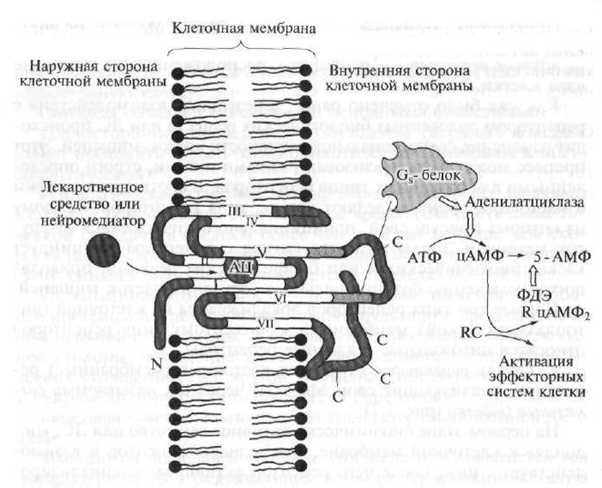
Fig. 1.8. Reprezentarea schematică a structurii „serpentinei”
receptor:
N este partea polipeptidică a receptorului situat deasupra membranei celulare; C este partea polipeptidică a receptorului situat sub membrana celulară; AC - centrul activ al receptorului cu care medicamentul interacționează; merge neurotransmițător; ATP - adenozina trifosfat - mesager secundar; cAMP - monofosfat de adenozină ciclică; 5-AMP - adenozin-5 "monofosfat; PDE - fosfodiesterază; R. RC - enzimă dependentă de cAMP (proteina kinaza) cu subunități regulatoare și catalitice (care accelerează reacția); 1-VII - lanțuri polipeptidice ale receptorului serpentin
Receptorii serpentinici includ lanțuri polipeptidice convolute (un polipeptid este un compus cu greutate moleculară mare, care este un lanț de resturi de aminoacizi interconectate) care pătrund membrana celulară de șapte ori.
Substanțele sau medicamentele biologice active endogene se pot lega de așa-numitul „buzunar” format de lanțul polipeptidic și localizat în grosimea membranei celulare, ceea ce implică formarea unui semnal de activare, care este transmis părților din lanțul receptor situat în citoplasma celulei. Proteinele de semnal G interacționează cu cy-
secțiuni stolice (intracelulare) ale podelei și lanțului peptidic; activează și lansează o cascadă de reacții biochimice care își schimbă activitatea funcțională despre celula țintă, adică. inițiază un răspuns farmacologic primar.
În prezent, sunt cunoscute mai multe tipuri de proteine \u200b\u200bG semnal.
Semnal G, -proteine. Aceste proteine \u200b\u200bde semnalizare, de regulă, activează un element efector - enzima adenilat ciclază, care la rândul său stimulează sinteza unui al doilea mesager din celulă (de la ATP) - adenosină monofosfat ciclic (cAMP). Rolul biologic al cAMP ca mesager secundar este foarte important. De exemplu, o creștere a conținutului său în celulele inimii atrage o creștere a frecvenței și a rezistenței contracțiilor inimii. În plus, o creștere a concentrației de cAMP în diferite celule țintă determină relaxarea mușchilor netezi ai vaselor și bronhiilor, mobilizarea rezervelor de energie (descompunerea carbohidraților din ficat), inhibă capacitatea de agregare a trombocitelor, scade tonul miometrului (mușchiului uterin) și vezicii urinare etc.
O serie de neurotransmițători, cum ar fi adrenalina (prin activarea receptorilor β-adrenergici), dopamina (prin activarea receptorilor D 1 -dopaminei), adenozină (prin activarea receptorilor adenozinei A 2), aparțin substanțelor endogene biologic active, cu capacitatea de a activa proteinele de semnalizare Gs. histamina (prin activarea histaminei G 2 -reneptorop), serotonină (prin activarea receptorilor serotoninei 5-HT 4), precum și a unui număr de hormoni, de exemplu, vasopresină (prin stimularea receptorilor V 2 -vasopresinei) etc.
Semnal G i -proteine. Spre deosebire de proteinele semnalului G, activarea proteinelor semnalului G i nu stimulează, ci inhibă activitatea elementului efector, enzima adenilat ciclază, care implică o scădere a concentrației de cAMP în celulele țintă ale mesagerului secundar. O scădere a conținutului de cAMP în celulele țintă determină o scădere a contracțiilor cardiace, o creștere a tonusului vaselor de sânge și a bronhiilor, adică. efectul opus creșterii conținutului de cAMP asupra celulelor țintă. În plus, o serie de proteine \u200b\u200bG i de semnalizare sunt implicate în reglarea activității funcționale a canalelor ionice transmembrane Ca 2+ și K +.
O serie de neurotransmițători, de exemplu, adrenalină și norepinefrină (prin activarea unui 2-adrenoreceptori), dopamină (prin activarea receptorilor D 2 - dopamina), adenozină (prin activarea A 1 receptori adenozinici), acetilcolină (prin activarea receptorilor muscarinici M 2 și M 4) etc.
Proteine \u200b\u200bde semnal G ^. Aceste proteine \u200b\u200bde semnalizare contribuie la activarea unui alt element efector al celulelor țintă, enzima fosforilazică C, care la rândul său stimulează formarea de mesageri secundari, diacilglicerol (DAG) și inozitol-1,4,5-trifosfat (ITP) în celulele țintă. Primul dintre ele (DAG) este asociat cu membrana celulară și inițiază reacții biochimice implicate în reglarea stării contractile, creșterea și divizarea celulelor și secreția anumitor hormoni de către celulele țintă. Sub influența enzimei fosfolipazei A 2, DAG poate fi metabolizat în acid arahidonic, care ia parte la sinteza substanțelor biologic active, cum ar fi eicosanoizii - prostaglandini, prostaciclini, tromboxani, leucotriene (vezi T. I, p. 478).
Al doilea mesager secundar, ITF, nu este fixat pe membrana celulară și se deplasează în mediul intracelular (citosol), unde inițiază eliberarea ionilor de Ca 2+ din depozitele de celule, adică. promovează tranziția ionilor de Ca 2+ inactivi la forma activă.
Mulți cercetători consideră ioni Ca 2+ ca un mesager terțiar sau un intermediar. Acest lucru se datorează faptului că rolul ionilor de Ca 2+ în reglarea activității funcționale a celulelor este foarte important. Ionii de Ca 2+ pot intra în celulă din mediul extern prin canale ionice speciale transmembranare și / sau eliberate din depozitele de celule. Principalul depozit (locul de acumulare de ioni Ca 2+ inactivi) în celulă este reticulul endoplasmatic sau sarcoplasmic (reticulum sarcoplasmaticunr, sinonim: reticulul endoplasmic - organelul intracelular, care este un sistem de tubule și cisterne situate în citoplasmă, care este limitată de furnizarea membranei; transportul substanțelor în citoplasmă). Ionii Ca (2) liberi (activi) care provin din reticulul sarcoplasmic la citoplasmă interacționează cu unele proteine \u200b\u200bde legare a Ca 2+, dintre care cea mai importantă este calmodulina. Complexul „calmodulin-Ca 2+” și / sau complexele de ioni Ca 2+ cu alte proteine \u200b\u200bcare leagă calciul declanșează o cascadă de reacții biochimice în celulă. Drept urmare, în funcție de organele țintă în care se produce acest proces, se inițiază o creștere a funcției contractile a miocardului și a mușchiului scheletului, o creștere a tonusului mușchilor netezi ai vaselor de sânge, bronhiilor și uterului, o creștere a activității secretorii a țesutului glandular, stimularea eliberării neurotransmițătorilor din terminațiile nervoase etc. . S-a dovedit, de asemenea, că ionii de Ca 2+ au capacitatea de a crește activitatea enzimelor implicate în metabolismul proteinelor, carbohidraților și grăsimilor.
În plus față de relația directă între mesageri secundari - DAG și ITF și, prin urmare, semnale proteine \u200b\u200bG q, ionii Ca 2+ în condiții fiziologice au o interacțiune destul de complexă cu mesagerul secundar cAMP, a cărei activitate este reglată de semnalul G s și proteinele G i. Astfel, s-a arătat că ionii de Ca 2+ liberi care intră în citoplasma unei celule nervoase prin sistemul calmodulin-Ca 2+ inițiază o scădere a conținutului de iAMP în celulă. În același timp, pentru menținerea stării deschise a canalelor de ioni de calciu în celulă, sunt necesare concentrații mari de cAMP, adică. scăderea conținutului de cAMP inițiat de complexul calmodulin-Ca 2+ atrage încetarea aportului de ioni Ca2+ liberi în citoplasmă. Pe de altă parte, există dovezi că mesagerul secundar cAMP îmbunătățește absorbția ionilor Ca2+ liberi de către reticulul sarcoplasmic, adică. promovează tranziția ionilor de Ca 2+ de la forma liberă, activă, la forma inactivă legată.
Ca urmare a creșterii conținutului de mesageri secundari în celulele țintă - DAG și ITF - tonusul mușchilor netede crește, secreția de glande crește, eliberarea de neurotransmițători din terminațiile presinaptice este facilitată, capacitatea de agregare a trombocitelor etc.
Substanțele active biologic active, cu capacitatea de a activa proteinele semnalului C, includ neurotransmițători precum norepinefrină (prin activarea unui 1 -adrenoreceptori), acetilcolină (prin activarea receptorilor muscarinici M 1 și M 3), serotonină (datorită activării serotoninei Receptorii 5-HT 2a), histamina (datorită activării receptorilor histaminici H1), precum și a altor substanțe endogene active biologic, de exemplu, bradininina și angiotensina.
În prezent, pe lângă semnale G-proteine \u200b\u200benumerate (G s, G |, G q), au fost identificate alte proteine \u200b\u200bG semnal - G s, G i, G q, al căror rol fiziologic este încă complet neclar. Dar, în același timp, există dovezi că, de exemplu, semnalul C o proteină este implicat în reglarea activității funcționale a canalelor ionice transmembranare.
Unitatea funcțională a receptorilor de tip 11 este o proteină transmembrană (care pătrunde pe întreaga grosime a membranei celulare) proteină (enzimă). Receptorul în sine este format din două fragmente identice, care se numesc monomeri. Monomerii sunt localizați la o distanță nesemnificativă unul de celălalt și monomerul în sine este format din două subunități active funcțional - domenii, interconectate de un segment polipeptidic care intersectează membrana bicapa lipidică (Fig. 1.9). Subunitatea a monomerului iese deasupra suprafeței exterioare a membranei și este responsabilă pentru legarea receptorului la substanțe biologic active, iar subunitatea P este cufundată în citoplasma celulei.

Fig. 1.9. Structura receptorului de tip II (explicația din text): 1 - a-subunitate a monomerului; 2 - subunitatea β a monomerului
După legarea substanței biologic active de subunitatea α a receptorului, receptorul se schimbă de la o stare monomerică inactivă la o stare dimerică activă în care doi monomeri se combină în planul membranei (vezi Fig. 1.9). În acest caz, activitatea enzimatică a subunității β-citoplasmice a receptorului este stimulată, ca urmare, o cascada de reacții biochimice care își schimbă starea funcțională este lansată în celula țintă.
Ca o enzimă transmembrană care formează receptorul, de regulă, sunt utilizate astfel de enzime precum tirozin kinază sau guanilat ciclază.
Un exemplu de receptor al tirozin kinazei este receptorii insulinei (vezi T. 1, p. 435).
Calea de transmitere a semnalului guanilate ciclază începe cu interacțiunea subunității a receptorului cu o substanță endogenă biologic activă, de exemplu, cu factor natriuretic atrial (ANF), care este o substanță biologic activă secretată de celulele atriale și implicată în reglarea contracțiilor cardiace. Ca urmare a acestei interacțiuni, are loc o modificare a configurației receptorului, care constă în combinarea monomerilor săi într-un dimer. Acest proces activează partea enzimatică a receptorului situat în subunitatea sa citosolică, adică. enzima guanylate ciclază, care la rândul său promovează o creștere a concentrației de mesager secundar de guanidină ciclic-3,5 "monofosfat (cGMP) în celula țintă. O creștere a concentrației de cGMP în celulele țintă declanșează o cascadă de reacții biochimice care își schimbă starea funcțională, de exemplu, relaxarea celulelor musculare netede nave.
Receptorii de tip III includ receptori care, sub influența substanțelor biologice active biologice - neurotransmițători, asigură trecerea ionilor corespunzători prin membrana celulară, ceea ce duce la o schimbare a încărcării sale electrice (potențiale).
În structura lor, receptorii tina III reprezintă un canal care pătrunde în stratul lipidic al membranei celulare, format din mai multe unități polipete (Fig. 1.10). De exemplu, receptorul nicotinic (H) este un canal cu un diametru de 8 nm, format din cinci subunități polipeptidice (a - două, β, γ, d) (vezi Fig. 1.10). Când neurotransmițătorul acetilcolină interacționează cu o porțiune (domeniu) care iese deasupra suprafeței membranei celulare - subunitatea α a receptorului - structura acestuia se schimbă și se deschide un canal central, prin care ionii Na + intră în celula țintă în funcție de gradientul de concentrație, ceea ce implică o modificare a funcționalității sale activitate. Pe lângă receptorii H-colinergici, receptorii pentru acidul gamma-aminobutiric și aminoacizii excitatori aparțin receptorilor de tip III.
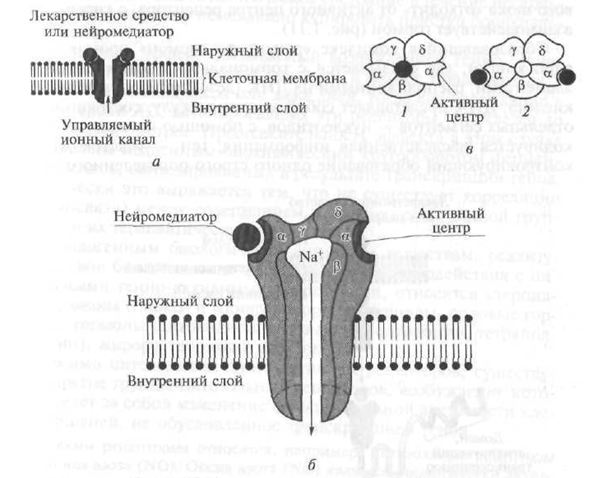
Fig. 1.10. Diagrama de structură a receptorului tip 111:
a este o diagramă de circuit; b - canal ionic transmembranar (în context); c - canal ionic transmembran (vedere de sus); / - canal în stare inactivă (închisă); 2 - canal în starea activă (deschisă); a. Subunitățile polipeptidice β, γ, d - canal
Receptorii de tip IV includ receptorii intracelulari și nucleari. Substanțele active biologic care interacționează cu acest tip de receptori sunt compuși lipofili (ușor solubili în grăsimi), prin urmare, ei pătrund cu ușurință în membrana celulară și ajung la receptorii lor intracelulari. Receptorii intracelulari includ receptorii pentru hormoni, precum și alte substanțe biologic active.
Mecanismul de interacțiune a hormonilor cu receptorii intracelulari este destul de complicat, însă poate fi reprezentat schematic după cum urmează. Prin structură, receptorul intracelular pentru hormoni este un polipeptid format din mai multe unități funcționale - domenii. În absența hormonului, receptorul este inactiv datorită faptului că centrul activ este blocat de o proteină specializată - așa-numita proteină de șoc termic. În cazul în care hormonul „se apropie” de receptor, proteina de șoc termic „se îndepărtează” de centrul activ al receptorului, cu care hormonul interacționează (Fig. 1.11).
Complexul receptor-hormonal rezultat pătrunde în nucleul celulei, unde se leagă de elemente sensibile la hormoni, localizate pe ADN (acid dezoxiribonucleic; ADN-ul este o macromoleculă formată din segmente individuale - nucleotide, cu ajutorul cărora informațiile ereditare sunt codificate în gene; gena - o bucată de ADN care controlează formarea unei proteine \u200b\u200bstrict definite
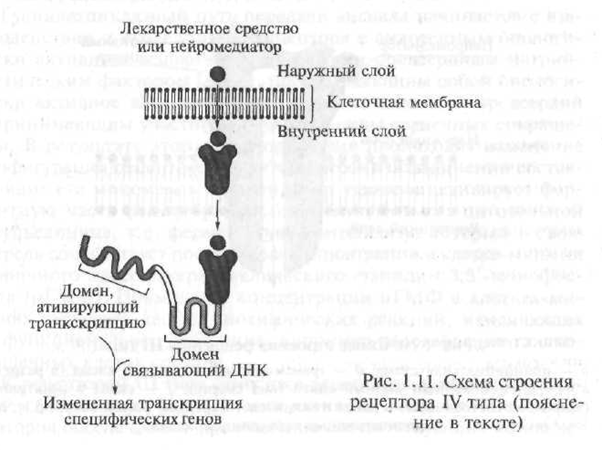
Fig. 1.11. Schema structurii receptorului de tip IV (explicație în text)
ka). Ca urmare a acestei interacțiuni, procesul de transcriere a genelor este început - procesul de transfer al informațiilor conținute în codul genetic de la molecula ADN la molecula de ARN de informație (ARNm, sin: matrică ARN - ARNm). Transcrierea este primul pas în formarea proteinelor într-o celulă. ARNm rezultat. părăsește nucleul celular și se deplasează spre ribozomi - organele intracelulare responsabile de sinteza proteinei din celulă. În literatura medicală specială, receptorii, a căror activare determină procesul de transcriere a genelor, sunt denumiți receptori activi genetic.
De regulă, răspunsul celulelor țintă la excitația receptorilor activi genici se dezvoltă destul de lent, ceea ce are o importanță clinică foarte importantă.
În primul rând, răspunsul celulelor țintă este întârziat în timp, deoarece necesită sinteza de noi proteine, care durează de obicei 20-30 de minute, adică. hormonii, activând receptorii de tip IV, nu sunt capabili să schimbe starea patologică în câteva minute, de exemplu, opresc imediat un atac de astm bronșic.
În al doilea rând, efectul cauzat de excitația receptorilor activi genici este destul de lung și poate dura câteva ore sau chiar zile, în timp ce conținutul din plasma sanguină a medicamentelor care au activat acești receptori scade la zero mult mai rapid. Durata efectului în acest caz se datorează circulației biochimice lente a enzimelor și proteinelor sintetizate ca urmare a transcrierii genelor. Clinic, acest lucru este exprimat prin faptul că nu există nicio corelație (relație) între conținutul plasmatic al unui anumit grup de medicamente și efectul lor terapeutic.
Substanțele active biologic endogene care își realizează efectele biologice prin interacțiunea cu receptorii citosolici activi ai genelor includ hormoni steroizi (gluco- și mineralocorticosteroizi, hormoni sexuali), hormoni tiroidieni (triiodotironină, tetraiodotironină) și vitamina D solubilă în grăsimi.
În plus față de receptorii citosolici activi ai genelor, există și alte grupuri de receptori citosolici, a căror excitație implică o schimbare a activității funcționale a celulelor țintă, nu datorită transcrierii genelor.
Astfel de receptori includ, de exemplu, receptori citosolici pentru oxidul nitric (N0). Oxidul nitric (N0) este o substanță activă biologic formată în endoteliul vascular. Ca substanță biologică activă endogenă, oxidul nitric a fost izolat pentru prima dată din porturile iepurelui de către fiziologul american R.F. Furchgott în 1987 și a fost denumit „factor relaxant endotelial - ORF”. Oxidul nitric este un compus lipofil care pătrunde cu ușurință în membrana celulară, unde interacționează cu receptorii citosolici specifici, ceea ce implică activarea enzimei guanylate ciclazei. Acesta din urmă, la rândul său, stimulează sinteza mesagerului secundar cGMP, care declanșează o cascadă de reacții biochimice intracelulare care duc la relaxarea celulelor țintă, a celulelor musculare netede vasculare.
Astfel, în prezent, se disting patru mecanisme principale și, în consecință, tipurile de receptori IV, datorită interacțiunii cu care substanțele endogene active biologic și / sau analogii lor sintetici, adică. Medicamentele pot afecta starea funcțională a celulelor țintă.
Cu toate acestea, acest lucru nu înseamnă că numărul de receptori cunoscuți pentru substanțele biologic active este limitat la 4. Sunt imensurabil mai mari. Acest lucru se datorează faptului că, prin același mecanism fundamental de acțiune, un număr foarte mare de substanțe biologice endogene din diferite structuri chimice pot influența activitatea funcțională a celulelor. De exemplu, neurotransmițătorii norepinefrină și histamină, care sunt diferite în structura lor chimică și, prin urmare, în receptorii cu care interacționează, transmit un semnal excitator către celulele țintă prin același mecanism de bază - stimularea activității proteinelor semnal G, adică. ambele interacționează cu receptorii de tip I.
Prin urmare, toți receptorii cunoscuți în prezent sunt clasificați nu numai pe caracteristicile transmiterii semnalului către structurile intracelulare ale celulelor țintă, ci și pe numele acelor substanțe endogene active biologic cu care interacționează în mod specific.
Trebuie menționat că receptorii și-au primit numele ținând cont de numele substanțelor biologice active biologice endogene cu care interacționează, cu mult înainte ca mecanismele de transmitere a semnalului către celulele țintă să devină cunoscute.
Substanțele active biologic active care își realizează efectele prin interacțiunea cu receptorii lor specifici includ neurotransmițători (acetilcolină, norepinefrină, dopamină, histamină, serotonină etc.), hormoni, substanțe biologic active de origine tisulară - autocaide (prostaglandine, tromboxane, leucotriene, etc. bradikinină, angiotensină etc.). În literatura medicală specială, toate aceste substanțe sunt adesea combinate sub termenul „liganzi” (din Lat. Ligo - pentru a se lega, adică o substanță capabilă să se lege cu receptorul).
Astfel, receptorii își iau numele de la numele liganzilor lor specifici. De exemplu, receptorii pentru neurotransmițătorul dopaminei se numesc dopamină, hormon insulinic - insulină, leucotriene augocaid - leucotriene etc.
În viitor, în textul manualului, pentru a evita confuziile în ceea ce privește mecanismul de transmitere a semnalului de la receptor la formațiuni intracelulare, va fi utilizat termenul „tip de receptor”, iar când vine vorba de numele receptorului din cauza ligandului care interacționează cu acesta , se va folosi termenul „specie receptor”.
De regulă, mulți receptori ai aceleiași specii sunt împărțiți în mai multe subtipuri, de exemplu, receptorii adrenergici sunt împărțiți în receptorii a și β-adrenergici, receptorii colinergici în receptorii M- și N-colinergici etc. În cele mai multe cazuri, subspecii sunt, de asemenea, subdivizate în grupuri mai mici: β 1 - și β 2 -adrenoreceptori, N n - și N m-receptori colinergici etc.
Identificarea sub-speciilor de receptori și studiul mecanismelor prin care substanțele endogene active biologic interacționează cu aceștia sunt foarte importante pentru farmacologia modernă, deoarece permite crearea de medicamente care interacționează cu o subspecie strict definită a receptorului. Deci, de exemplu, divizarea receptorilor β-adrenergici în β 1 (localizată în principal pe membrana celulară a celulelor inimii) și β 2 (localizată, de exemplu, pe membranele celulare ale celulelor musculare netede ale bronhiilor), a făcut posibilă crearea de medicamente care afectează selectiv mușchiul cardiac (β 1 -adrenostimulante ) - medicamentul nonachlazină și afectează selectiv mușchii netezi ai bronhiilor (β 2 -adrenostimulante) - medicamentul salbutamol etc.
Trebuie menționat că organele și țesuturile corpului nu conțin un număr constant de receptori și / sau subspecii lor, adică. este variabil. Atât procesele patologice, cât și medicamentele pot schimba numărul de receptori dintr-un organ.
De exemplu, boala coronariană este însoțită de o creștere a numărului de receptori 3-adrenergici din mușchiul inimii, iar la pacienții cu hipertensiune arterială, numărul receptorilor a și a β-adrenergici crește. Imipramina antidepresivă, cu utilizare prelungită, reduce cantitatea de receptori β-adrenergici din țesutul creierului. Există destul de multe astfel de exemple.
Afinitatea neurotransmițătorilor endogeni (produși în organism) sau a medicamentelor pentru receptori este caracterizată prin termenul „afinitate”, iar viteza și rezistența legării lor la receptori este desemnată prin termenul „afinitate”.
În mod natural, interacțiunea medicamentelor cu receptorul nu este un scop în sine, ci ar trebui să conducă la o anumită modificare a activității organelor sau țesuturilor corpului.
O astfel de schimbare, sau reacție, corespunzătoare semnificației funcționale a acestui receptor, se numește activitate internă a medicamentelor.
Medicamentele cu activitate intrinsecă și afinitatea receptorilor sunt agoniști, adică. acționează ca substanțe biologice endogene active.
De exemplu, fenilefrina, un stimulator al receptorilor a-adrenergici, are un efect asupra arteriolelor similare neurotransmițătorului norepinefrină, adică. el este un agonist al receptorilor a-adrenergici. În literatura medicală specială, pe lângă termenul „agonist”, este folosit uneori termenul „stimulent al receptorilor” sau „mimetic”, de exemplu, adrenomimetic, adică. Medicamente care stimulează receptorii adrenergici.
Medicamentele care au afinitatea receptorului, dar inhibă interacțiunea cu agonistii exogeni și endogeni cu numele de receptori, sunt numiți antagoniști.
De exemplu, blocantul receptorului M-colinergic al atropinei interferează cu interacțiunea neurotransmițătorului acetilcolinei M-receptorul colinergic; blocant (receptorii J-adrenergici propranolol, blocând β 2 -adrenostructurile plămânilor, împiedică efectul stimulant al β 2 -adrenostimulator al salbutamolului, adică atropina și propranololul sunt antagoniști ai receptorilor corespunzători.
În literatura medicală de specialitate, pe lângă termenul „antagonist”, termenul „blocanți ai receptorilor” sau „litiști”, de exemplu, anticolinergici, adică. medicamente care blochează receptorii colinergici.
Schematic, interacțiunea receptorilor cu agoniști și antagoniști este prezentată în Fig. 1.12.
Agoniștii pot exercita atât direct, cât și indirect, adică. acțiune indirectă.

Fig. 1.12. Schema de interacțiune a receptorului cu agonistul (a) și antagonistul (6) cu receptorul (explicația din text)
De exemplu, un agonist al receptorilor opioizi - morfina medicamentului - își realizează efectele prin stimularea directă (mu) p-, (kappa) k- și (delta) 8-receptori opioizi, adică. exercită un efect direct de stimulare asupra lor, în timp ce efedrina simpatomimetică își realizează efectele celulare prin stimularea indirectă sau indirectă a și a receptorilor β-adrenergici: ajută la deplasarea neurotransmițătorului norepinefrină de la terminațiile nervoase presinaptice, inhibă recaptarea acesteia prin terminații nervoase și crește sensibilitatea a-și p- adrenoreceptori la norepinefrină și epinefrină și stimulează eliberarea de adrenalină din cortexul suprarenal. Astfel, efedrina își realizează efectele farmacologice nu direct, ci printr-un neurotransmițător, în timp ce nu interacționează direct cu receptorul.
Antagoniștii, precum și agoniștii receptorilor își pot realiza efectele farmacologice direct sau indirect, blocând receptorii corespunzători.
De exemplu, blocantul receptorului histaminic H3 gastric, ranitidina, interacționează direct cu receptorii histaminici H 2 gastrici, inhibă secreția bazală de acid clorhidric, inhibând interacțiunea cu receptorul endogen de neurotransmițător histaminic, adică. blochează direct receptorul, în timp ce reserpina simpatolitică, care intră în corp, la fel ca efedrina simpatomimistică „ejectează” neurotransmițătorul norepinefrină de la terminațiile presinaptice, blocând simultan sinteza de noi „porții” de norepinefrină, ceea ce duce la o epuizare rapidă a rezervelor de mediator în presinaptic. Drept urmare, se creează o stare atunci când receptorii a-adrenergici ai vaselor „nu funcționează” din lipsa stimulatorului lor fiziologic noradrenapină. Putem spune că reserpina simpatolitică provoacă o blocare funcțională și indirectă a receptorilor a-adrenergici.
Unele medicamente combină proprietățile agoniștilor și antagoniștilor, adică. în anumite condiții, aceiași receptori excită sau se blochează. În acele cazuri în care componenta stimulatoare prevalează în efectul farmacologic al medicamentului, ei spun că medicamentul este un agonist parțial sau parțial.
Mecanismul de acțiune al agoniștilor parțiali sau parțiali este acela că aceste medicamente, care se apropie de receptor, sunt fixate pe el. Cu toate acestea, datorită particularităților structurii lor chimice, acestea îl excită în așa fel încât să nu determine un răspuns complet, ci doar parțial de stimulare. În același timp, acestea interferează cu interacțiunea altor medicamente și / sau neurotransmițătorul corespunzător cu acest receptor.
De exemplu, nalorfina, care este utilizată pentru a trata un supradozaj de analgezice narcotice, este un agonist parțial. Nalorfinul, care „urcă” la receptorul opioid, deplasează morfina din legătura sa, adică oprește efectul agonistic al morfinei asupra receptorului. Contactând receptorul, nalorfina într-o mică măsură are un efect stimulant asupra acestuia, care nu este în măsură să suprime activitatea centrului respirator, adică. are un efect agonistic parțial asupra receptorului opioid, în paralel cu acesta, îl blochează pentru morfină, prezentând astfel un efect antagonist în ceea ce privește morfina. Dezavantajul nalorfinei este că, în caz de supradozaj, efectul stimulant al acestuia asupra receptorului este îmbunătățit și inhibă centrul respirator, precum și morfină.
În acele cazuri în care un efect de blocare prevalează în efectul farmacologic al medicamentelor, ei spun că medicamentul este un antagonist cu activitate proprie sau internă.
Particularitățile mecanismului de acțiune al antagoniștilor cu activitate proprie pot fi luate în considerare pe exemplul β 1 -adrenoblocator al acebutalolului. Acebutalolul aparține grupului de blocanți β-adrenergici cu activitate simpaticomimetică internă, adică. combină proprietățile unui blocant (litic) și un stimulent (mimetic) al β1 -adrenoreceptorilor.
Mecanismul de acțiune al acebutalolului se datorează particularităților structurii sale chimice, datorită căruia medicamentul, pe de o parte, blochează β1 -adrenoreceptorii miocardici, iar pe de altă parte, stimulează activitatea acestora, simulând astfel efectul fiziologic al mediatorilor - catecolaminele (vezi T. 1. p. 170).
Toate medicamentele în funcție de particularitățile acțiunii lor pot fi împărțite în două grupuri mari - cu efecte specifice și nespecifice.
Medicamentele care au un efect nespecific includ medicamente care au o gamă largă de activitate farmacologică și diverse puncte de aplicare a efectelor. Acest grup include vitamine, stimulenți biogeni, antioxidanți, adaptogeni etc.
Medicamentele cu efect specific includ medicamente caracterizate prin proprietățile agoniștilor sau antagoniștilor receptorilor corespunzători, adică. baza mecanismului lor de acțiune este capacitatea de a interacționa cu receptori specifici pentru aceștia.
Cu toate acestea, trebuie menționat că efectul farmacologic al medicamentelor care au un efect specific asupra receptorilor nu va fi întotdeauna foarte specializat, de exemplu, reducerea frecvenței cardiace (HR) sau suprimarea secreției de suc gastric. Efectul farmacologic total al multor medicamente va depinde de câte și în care sunt localizate organele și țesuturile formațiunilor receptorilor specifice pentru acest medicament.
De exemplu, receptorii β-adrenergici sunt localizați în mușchiul inimii, peretele vascular, bronhiile, uterul, țesutul adipos, mușchiul scheletului etc. Ca urmare a acestui fapt, medicamentele care stimulează receptorii β-adrenergici vor determina, într-o anumită măsură, creșterea forței și frecvenței contracțiilor inimii, expansiunea vaselor de sânge, bronhiile și tonul inferior al uterului, adică au un efect sistemic asupra organismului.
Efectul specific al medicamentelor poate fi selectiv sau selectiv și, în consecință, neselectiv sau neselectiv. Selectivitatea medicamentelor este determinată de faptul dacă afectează toate subspecii oricărui receptor sau subspecia lor specifică.
De exemplu, propranololul blocant β-adrenergic neselectiv blochează atât β 1 -adrenoreceptorii localizați în țesutul miocardic cât și β 2-adrenoreceptorii localizați, în special, în țesutul pulmonar, adică, au un efect non-selectiv (non-selectiv) asupra ambelor subspecii de receptori β-adrenergici. . în timp ce blocantul selectiv β1-adrenergic atenolol în doze terapeutice blochează numai receptorii adrenergici miocardici β 1, adică. exercită un efect selectiv (selectiv) asupra unui receptor β-adrenergic specific.
Clarificarea rolului fiziologic și localizarea subspeciilor receptor permite crearea de medicamente extrem de eficiente care acționează selectiv asupra diferitelor subspecii de receptori.
De exemplu, medicamentele care blochează receptorii β 1-adrenergici sunt utilizate pe scară largă în clinică pentru tratamentul bolilor coronariene, hipertensiunii arteriale și medicamentelor care stimulează receptorii β2-adrenergici și-au găsit aplicația pentru tratamentul astmului bronșic. Medicamentele care blochează receptorii de histamină H1 sunt de preferat prescrise în practica clinică pentru a preveni și / sau a aresta reacții alergiceîn timp ce medicamentele care blochează receptorii histaminici H 2 sunt eficienți în tratarea ulcerelor gastrice și duodenale.
Crearea medicamentelor cu efect selectiv a optimizat în mare măsură utilizarea clinică a acestora, deoarece acestea au redus semnificativ efect secundar.
Baza mecanismului de acțiune al medicamentelor, de regulă, este capacitatea lor de a iniția (declanșa) procese biochimice și / sau biofizice complexe care în final modifică și / sau optimizează activitatea funcțională a celulei țintă. Medicamentele își pot realiza efectul asupra organelor și / sau celule țintă prin: interacțiune chimică directă; interacțiune fizico-chimică pe membrana celulară; acțiuni asupra enzimelor de specialitate; acțiuni asupra genelor de reglementare; acțiuni asupra receptorilor specifici.
Interacțiunea chimică directă a medicamentelor.
Acest mecanism de acțiune al medicamentelor este destul de rar și poate fi realizat în afara celulei, de exemplu, în lumenul stomacului sau al intestinelor. Esența sa constă în faptul că medicamentele intră într-o reacție chimică directă cu molecule și / sau ioni care se formează în organism într-o stare normală sau patologică. Un exemplu de interacțiune chimică directă este reacția chimică de neutralizare a acidului clorhidric al stomacului atunci când luați medicamente antiacide.
Interacțiunea fizico-chimică a medicamentelor pe membrana celulară.
Una dintre principalele funcții ale membranei citoplasmatice este implementarea schimbului de ioni între citoplasmă și mediul extracelular. Schimbul de ioni transmembranari poate avea loc, de asemenea, prin canale ionice transmembrane speciale dependente de tensiune - sodiu, potasiu, calciu, clor etc. Unele medicamente, care ajung la membrana celulară, interacționează cu aceste canale și le modifică activitatea funcțională. Deci, de exemplu, acțiunea antiaritmică a medicamentului clasei IA - chinidină - se bazează pe capacitatea sa de a bloca trecerea ionilor de Na + prin canalele de sodiu transmembranare.
Efectul medicamentelor asupra enzimelor specializate.
O cantitate relativ mică de medicamente își realizează efectul farmacologic prin schimbarea activității unor enzime celulare specializate. Se numesc medicamente care cresc activitatea enzimelor celulare inductorienzime. O astfel de acțiune este deținută, de exemplu, de somnifere și de medicamentul anticonvulsivant fenobarbital, care îmbunătățește semnificativ activitatea enzimelor hepatice microsomale. Semnificația biologică a acestui efect al fenobarbitalului și al medicamentelor conexe vor fi discutate mai jos.
Se numesc medicamente care inhibă activitatea enzimelor specializate inhibitorienzime. Deci, de exemplu, un antidepresiv din grupul inhibitorilor monoaminoxidazei (MAO), medicamentul pirlindol își realizează efectul antidepresiv prin inhibarea activității enzimei MAO în sistemul nervos central. Capacitatea de a inhiba activitatea enzimei acetilcolinesterază stă la baza activității farmacologice a medicamentelor anticolinesterază, de exemplu fizostigmină. Se știe că în condiții fiziologice, acetilcolinesteraza inactivează (distruge) acetilcolina, un neurotransmițător care transmite excitația în sinapsele sistemului nervos parasimpatic. Physostigmine, care suprimă activitatea acetilcolinesterazei, contribuie la acumularea în sinapsele sistemului parasimpatic al neurotransmițătorului acetilcolină, ca urmare a creșterii tonusului sistemului nervos parasimpatic, care se manifestă la nivelul sistemului prin dezvoltarea bradicardiei, scăderea presiunii sanguine (BIT), creșterea tensiunii arteriale (BIT), elev etc. Medicamentele pot interacționa reversibil și ireversibil cu enzimele. De exemplu, medicamentul enalapril inhibă reversibil activitatea enzimei care convertește angiotensina, ceea ce implică, în special, o scădere a tensiunii arteriale, în timp ce substanțele toxice organofosforice inhibă ireversibil activitatea acetilcolinesterazei. Efectul medicamentelor asupra genelor de reglementare.În prezent, oamenii de știință fac încercări de a crea medicamente care să realizeze efectele lor farmacologice, afectând direct activitatea fiziologică a genelor reglatoare. Această tendință pare deosebit de promițătoare după ce structura genomului uman a fost descifrată în 2000. Se crede că normalizarea selectivă a funcției genelor reglatoare sub influența medicamentelor va face posibilă obținerea unui succes în tratamentul multor boli, inclusiv anterior incurabile. Efectul medicamentelor asupra receptorilor.Înainte de a trece la caracteristicile interacțiunii medicamentelor cu receptorii, este necesar să clarificăm ce înțelegem prin termenul „receptor” (din lat. recipio - ia, ia). De la cursul fiziologiei se știe că termenul „receptor” înseamnă formațiuni extrem de specializate care sunt capabile să perceapă, să transforme și să transmită energia unui semnal extern către sistemul nervos. Acești receptori sunt numiți senzorial(din lat. sensus - sentiment, senzație, percepție). Receptorii senzoriali includ receptorii organelor auzului, vederii, mirosului, gustului, atingerii etc. Receptorii senzoriali ai acestor organe aparțin așa-numitilor exteroreceptori. Dacă prezența organelor senzoriale care răspund la stimuli externi de iritație este cunoscută încă din cele mai vechi timpuri, atunci prezența receptorilor senzoriali în interiorul corpului a fost pusă la îndoială până la mijlocul secolului al XIX-lea. Pentru prima dată, fiziologul rus I.F. Sion, care a arătat în 1866 o scădere a tensiunii arteriale din cauza iritării aortice într-un experiment de iepure. Această descoperire a dat naștere căutării și studiului receptorilor localizați în interiorul corpului și acești receptori înșiși au fost numiți interoreceptori. Kînceputul secolului XX a fost dezvăluit un număr suficient de interoreceptori senzoriali și s-a dovedit rolul lor important în reglarea funcțiilor fiziologice ale organismului. În 1905, J. Langley a dovedit că atunci când un medicament este aplicat pe o membrană celulară, un efect farmacologic se dezvoltă dacă este aplicat doar pe o anumită zonă a acestuia. Mai mult, acest site nu constituie decât o mică parte din suprafața totală a suprafeței celulare. Această observație a permis lui J. Langley să concluzioneze că pe membrana celulară există site-uri de receptori specializați care interacționează cu medicamente. Cu toate acestea, prioritatea în crearea teoriei receptorilor a acțiunii medicamentelor aparține fiziologului german P. Ehrlich, care în 1906 a introdus pentru prima dată termenul de „receptor” și a formulat postulatul „medicamentul nu funcționează dacă nu este fixat pe membrana celulară”. Conform teoriei lui P. Ehrlich, o moleculă de medicament are două grupe active din punct de vedere funcțional, dintre care una asigură fixarea acesteia pe suprafața celulelor din regiunea receptorului de droguri, iar cea de-a doua grupă funcțională interacționează cu receptorul și declanșează un lanț complex de reacții biochimice care îi modifică activitatea fiziologică (celulară). . Astfel, încă de la începutul secolului XX. a devenit evident că există cel puțin două clase de interoreceptori: receptorii senzorialitransmiterea informațiilor despre starea organelor interne și a țesuturilor corpului din sistemul nervos central; receptorii celularicare interacționează cu medicamente care modifică activitatea funcțională a celulelor țintă.<>Imediat, trebuie menționat că, în viitor, în textul manualului, pentru a evita confuziile în terminologie, receptorii pentru medicamente și substanțele biologic active, adică. celular, sau citoreceptorriu,va fi notat de termenul „receptor”, în timp ce interoreceptorii senzoriali vor fi denotați de un termen care caracterizează activitatea lor funcțională, de exemplu, „baroreceptori”, „receptori ai durerii” etc. Descoperirea făcută de P. Ehrlich pe membrana celulară a receptorilor medicamentelor a servit ca punct de plecare pentru dezvoltarea științei farmacologice, în special farmacodinamica, una dintre principalele sarcini ale căreia este studierea mecanismelor receptorului de acțiune a medicamentelor. În prezent, structura unui număr mare de receptori celulari, a fost dezvăluită caracteristicile interacțiunii anumitor compuși activi biologic cu aceștia, ceea ce a făcut posibilă, pe de o parte, înțelegerea mecanismului de acțiune a medicamentelor cunoscute, iar pe de altă parte, a fost baza pentru crearea de noi medicamente extrem de eficiente. În mod natural, este greu de imaginat că în cursul evoluției, în corpul uman s-au format receptori pentru diverse medicamente sintetice (obținute chimic), mai ales că marea majoritate a medicamentelor prezentate pe piața farmaceutică modernă au fost sintetizate în ultimii 50 de ani sau mai puțin. Este dovedit că aparatul receptor al celulei este o formație funcțional-structurală foarte veche. Deci, a- și b-adrenoreceptori (receptori, a căror interacțiune norepinefrină și adrenalină afectează activitatea funcțională a celulei) se găsesc nu numai în celulele animale, ci și pe membranele celulare ale celulelor plantei, de exemplu, în celulele nittellei plantelor, unde a- și b- adrenoreceptorii reglează mișcarea protoplasmei (conținutul celulelor). Atunci care sunt receptorii pentru medicamentele descoperite de P. Ehrlich și de ce interacționează cu aceștia? În prezent, nu există nicio îndoială că așa-numiții receptori de droguri sunt de fapt receptori pentru substanțele biologice active endogene (produse în organism) implicate în reglarea activității funcționale a organelor interne și a țesuturilor corpului. Astfel de compuși activi biologic includ substanțe eliberate de terminațiile nervoase la momentul transmiterii semnalului nervos, precum și hormoni, vitamine, aminoacizi etc. Pentru fiecare substanță biologică activă endogenă, există receptori strict specifici pentru aceasta. Deci, de exemplu, substanța activă biologic produsă în organism, adrenalina, poate activa strict specific - și receptorii b-adrenergici și glucocorticosteroizii - hormoni ai cortexului suprarenal - interacționează numai cu receptorii glucocorticosteroizi strict specifici acestora. Medicamentele sintetice care își realizează efectele prin interacțiunea cu aparatul receptor al celulei, în structura lor chimică, sunt mai mult sau mai puțin similare cu compușii activi biologic endogeni care interacționează cu receptorii similari. De exemplu, medicamentele sintetice cu vasoconstrictor (care provoacă vasoconstricție), fenilefrina este apropiată în structura sa chimică de substanța endogenă biologic activă norepinefrină, prin urmare, la fel ca norepinefrina, are capacitatea de a stimula a-adrenoreceptorii. Uneori, datorită particularităților structurii lor chimice, medicamentele pot interacționa nu cu receptorul în sine, ci cu porțiunea adiacentă a membranei celulare. Deoarece în acest caz, medicamentul interacționează nu cu receptorul în sine, ci cu porțiunea adiacentă a membranei celulare, acestea nu vorbesc despre un efect excitant sau de blocare asupra receptorului, ci allostemetric(din greacă allos - alt efect, diferit). Ca urmare, poate apărea o modificare atât a structurii membranei adiacente receptorului, cât și a componentelor individuale ale receptorului în sine, ceea ce poate atrage o schimbare a sensibilității receptorului față de o substanță activă biologic specifică acestuia. În cazurile în care sensibilitatea receptorului la o substanță activă biologic crește, vorbiți despre sensibilizare(din lat. sensus - sentiment) sau despre sensibilizare(din lat. sensibilis - sensibilitate) a receptorului și în cazurile în care sensibilitatea receptorului scade, vorbim despre desensibilizareareceptor. Particularitatea efectului alosteric constă în faptul că medicamentele care au acest tip de mecanism de acțiune nu afectează în mod direct transmiterea unui impuls nervos, ci îl modifică în direcția dorită. De exemplu, mecanismul de acțiune al anxioliticelor (medicamente anti-anxietate; sinonime: tranchilizante), în structura lor chimică fiind derivați de benzodiazepină, se bazează pe fenomenul excitării alosterice a receptorilor postsinaptici de benzodiazepină. Excitația acestuia din urmă, la rândul său, promovează activarea receptorilor inhibitori postsinaptici ai acidului gamma-aminobutiric (receptorii GABA), care se manifestă clinic prin eliminarea simptomelor bolilor nevrotice precum anxietatea, anxietatea, frica etc. Receptorii, care interacționează cu care, o substanță sau un medicament biologic activ schimbă în orice mod starea funcțională a celulei țintă, sunt numiți specifice. Pe lângă receptorii specifici, așa-numitele nespecificepentru receptorii medicamentelor. În literatura medicală de specialitate, acești receptori mai sunt numiți „locul pierderii” de medicamente. Prin contactarea unor astfel de receptori, medicamentele nu au niciun efect biologic, dar ele însele devin biologice inactive. Un exemplu de acest tip de receptori poate servi ca receptori localizați pe proteine \u200b\u200bplasmatice, în special, pe proteine \u200b\u200bsolubile în apă - albumina. Structura receptorilor este destul de complexă, dar majoritatea sunt macromolecule proteice sau glicoproteine, care pot include și ioni, lipide, acizi nucleici etc. Receptor i.e. macromolecula de proteine \u200b\u200bcare o formează se caracterizează printr-o aranjare spațială specifică pentru fiecare receptor, a grupelor sale chimice. Macromolecula proteică care formează receptorul poate fi integrată (cufundată) în stratul lipidic al membranei citoplasmatice sau localizată în interiorul celulei. Principala funcție a unui receptor celular este „recunoașterea” unui semnal chimic transmis acestuia printr-o substanță endogenă biologic activă și / sau medicamente și transformarea acestuia într-un răspuns celular biochimic și / sau biofizic corespunzător. Se credea anterior că medicamentele sau substanțele endogene active biologic interacționează cu receptorii de tip „cheie și blocare”, adică. receptorul are o astfel de structură care permite medicamentului să-ți găsească receptorul „tău”, să se conecteze la acesta și, așa cum era, „pornește” și „oprește-l”. Cu toate acestea, odată cu dezvoltarea științei medicale, a devenit evident că acest lucru nu este în întregime adevărat. În prezent, procesele moleculare de conversie a semnalelor extracelulare în celule intracelulare, care reglează funcția celulelor, au fost deja studiate destul de bine. mecanisme care au ca efect efectul interacțiunii substanțelor endogene active sau a medicamentelor cu receptorii. Atunci când interacționează cu receptorul unei substanțe endogene biologic active și / sau a unui medicament activ ca acesta apare conformație- modificarea spațială sub forma unei macromolecule proteice, care este declanșatorul pentru diverse procese intracelulare care determină răspunsul unei celule țintă la un mediator și / sau un medicament. De exemplu, activarea b 2 -adrenoreceptorilor mușchiului neted bronșic sub influența unui fenoterol b-adrenostimulator conduce la o creștere a activității enzimei adenilat ciclază, ceea ce contribuie la acumularea de adenozin monofosfat ciclic (cAMP) în celulă și, ca urmare, relaxarea celulelor. În termeni biologici generali, receptorii celulari pot fi considerați ca „organe senzoriale” strict specializate ale celulelor, prin intermediul cărora percep „informații” care provin, de exemplu, din sistemul nervos central și / sau din sistemul endocrin. În ciuda rolului important al aparatului receptor, receptorii ocupă doar o parte nesemnificativă a membranei celulare. De exemplu, celula receptorului M-colinergic nu ocupă cel mult 1/6 000 din suprafața sa. Studierea caracteristicilor interacțiunii medicamentelor cu receptorul, pe de o parte, ne permite să înțelegem baza mecanismului molecular al acțiunii sale, iar pe de altă parte, oferă informații despre ce schimbări ar trebui făcute în structura medicamentelor pentru a spori capacitatea acestuia de a interacționa cu acest receptor, adică. permite sinteza țintită de noi medicamente extrem de eficiente. În condiții fiziologice, diferiți receptori celulari nu funcționează independent, ci sunt într-o interacțiune constantă între ei, reglând astfel activitatea specifică a celulei. De exemplu, activarea receptorilor b-adrenergici cardiaci prin norepinefrină endogenă determină, în special, o creștere a numărului de contracții cardiace și activarea receptorilor M-colinergici ai celulelor cardiace de către acetilcolina endogenă, determină, dimpotrivă, o scădere a numărului de contracții cardiace. O mare contribuție la înțelegerea mecanismelor receptorilor de acțiune a medicamentelor a fost realizată prin descoperirea receptorilor pre și post-sinaptici. Bdiurn(din greacă sinopsis - conexiune) este o zonă de contact specializată între celulele nervoase sau alte structuri excitabile ale corpului, care asigură transmiterea informațiilor de intrare și păstrarea semnificației sale informaționale. Studiul structurii și rolului funcțional al sinapselor a fost început la sfârșitul secolului XIX. după ce histologul spaniol S. Ramon-i-Cajal (S. Ramon la Cajal) a sugerat prezența unui sistem de transmisie specializat în sistemul nervos central. Sinapsele și-au primit numele în 1897, când fiziologul englez Ch. Sherrington a propus acest termen să se refere la zona de contact dintre celulele nervoase. În prezent, există trei tipuri de sinapse: 1) sinapsele „electrice”, în care informațiile sunt transmise prin transferul unui semnal electric dintr-o membrană pre-sinaptică. Acest tip de sinapsă se numește efaps(din greacă ephapsis - contact strâns); 2) sinapsele „chimice” în care informațiile sunt transmise prin substanțe biologice speciale active - neurotransmitatori(din greacă neuron - nerv si lat. mediator - mediator; sinonime: mediator); 3) sinapsele „mixte” în care informațiile sunt transmise atât chimic cât și electric. Efectele farmacologice ale majorității medicamentelor care afectează funcțiile sinapselor sunt realizate prin efectul lor asupra unei etape particulare de transmitere a semnalului în sinapsele chimice, adică. în sinapsele de al doilea fel. De regulă, sinapsele chimice sunt clasificate de neurotransmițătorii care transmit impulsuri nervoase în ele, astfel: sinapsele în care acetilcolina acționează ca mediator sunt numite colinergice; sinapsele în care adrenalina și norepinefrina acționează ca mediator sunt numite adrenergice; sinapsele în care ATP și adenozina acționează ca mediator sunt numite purinergice; sinapsele în care acidul gamma-aminobutiric acționează ca mediator se numesc GABA-ergic etc. În prezent, structura sinapselor este bine înțeleasă. Sinapsa constă dintr-un proces presinaptic al unei celule nervoase (capătul axonului) și un aparat de recepție „semnal” situat pe membrana unei celule efectoare („executive”).
Farmacodinamică - efecte farmacologice, mecanisme de acțiune, localizarea acțiunii, tipuri de acțiune a medicamentelor.
Efecte farmacologice substanță medicinală - modificări ale activității organelor, sistemelor corporale care provoacă această substanță (de exemplu, creșterea contracțiilor cardiace, scăderea tensiunii arteriale, stimularea activității mintale, eliminarea fricii și tensiunii etc.).
Fiecare substanță provoacă o serie de efecte farmacologice caracteristice acesteia. În fiecare caz, sunt utilizate doar anumite efecte ale substanței medicamentoase, care sunt definite ca fiind principalul efecte. Efectele farmacologice rămase (neutilizate, nedorite) sunt denumite advers.
Mecanisme de acțiune medicamente - modalitățile în care substanțele provoacă efecte farmacologice sunt foarte diverse. Principalele opțiuni pentru mecanisme de acțiune includ acțiuni asupra:
- receptori specifici;
- enzime;
- canale ionice;
- sisteme de transport.
Majoritatea medicamentelor acționează receptori specifici. Acești receptori sunt reprezentați cel mai adesea de molecule proteice active funcțional, interacțiunea cu care dă naștere la reacții biochimice care duc la efecte farmacologice.
Există receptori specifici asociați cu membranele celulare (membrană) și receptorii intracelulari (citoplasmatic, nuclear).
Receptorii membranei (receptorii membranei citoplasmatice) se împart în:
- receptori direct cuplati la canalele ionice;
- receptori conjugați direct cu enzime;
- receptori care interacționează cu proteinele G.
K receptorii cuplati direct cu canalele ionice, includ, în special, receptorii Ν-colinergici și receptorii GABA A.
Când sunt stimulați receptorii colinergici Ν (receptorii colinergici sensibili la nicotină), canalele de sodiu cuplate direct cu acestea se deschid. Stimularea receptorilor olin-colinergici duce la descoperirea canalelor Na +, intrarea ionilor Na + în celulă, depolarizarea membranei celulare și un efect excitant.
Receptorii GABA A sunt cuplati direct cu canalele de clor. Stimularea receptorilor GABA A duce la descoperirea canalelor Cl, intrarea ionilor Cl, hiperpolarizarea membranei celulare și efectul inhibitor.
K receptori care sunt direct legați de enzimeinclud, în special, receptorii de insulină conjugați direct cu tirozin kinază.
Receptorii proteinei G - Receptori M-colinergici (receptori colinergici sensibili la muscarinici), receptori adrenergici, receptori dopaminici, receptori opioizi etc.
Proteinele G, adică proteinele care leagă GTP, sunt localizate în membrana celulară și constau din subunități α-, β- și γ. Când un medicament interacționează cu un receptor, subunitatea α a proteinei G se leagă de GTP (GTP) și acționează asupra enzimelor sau canalelor ionice.
Un receptor interacționează cu mai multe proteine \u200b\u200bG și fiecare complex al subunității α a proteinei G cu GTP acționează asupra mai multor molecule enzimatice sau asupra mai multor canale ionice. Astfel, mecanismul unui amplificator (amplificator) este implementat: atunci când un receptor este activat, activitatea multor molecule enzimatice sau a multor canale ionice se schimbă.
Unul dintre primii care au fost găsiți au fost proteinele G asociate cu β1 -adrenoreceptorii inimii. Atunci când inactivarea simpatică a inimii este activată, β1 -adrenoreceptorii sunt excitați; adenilat ciclaza este activat prin proteine \u200b\u200bG; cAMP este format din ATP, proteina kinazei este activată, sub acțiunea căreia canalele Ca 2+ sunt fosforilate și deschise.
O creștere a aportului de ioni Ca 2+ în celulele nodului sinoatrial accelerează a patra fază a potențialului de acțiune, frecvența impulsurilor generate crește - contracțiile inimii devin mai frecvente.
Descoperirea canalelor de Ca 2+ în fibrele miocardului de lucru duce la o creștere a concentrației de Ca 2+ în citoplasmă (aportul de Ca 2+ favorizează eliberarea de Ca 2+ din reticulul sarcoplasmic). Ionii de Ca 2+ se leagă de troponina C (o componentă a troponinei-tropomiozinei); Astfel, efectul inhibitor al troponinei-tropomiozinei asupra interacțiunii actinei și miozinei scade - contracțiile inimii sunt intensificate (Fig. 10).
Fig. 10. Mecanismul de creștere și întărire a contracțiilor cardiace în timpul stimulării β 1 -adrenoreceptori. AC - adenilat ciclază; PC - proteina kinaza; CA - nod sinoatrial; TTM - troponină-tropomiozină.
Când este activată inervația parasimpatică a inimii (nervii vagi), receptorii M 2-colinergici sunt excitați, iar prin proteine \u200b\u200bG, inhibarea ciclenei adenilate este slăbită - contracțiile cardiace sunt slăbite și slăbite (în principal contracțiile atriale sunt slăbite, deoarece inervația parasimpatică a ventriculelor este relativ slabă).
Astfel, proteinele G pot avea un efect stimulator și inhibitor asupra adenilat ciclazei. Proteinele G stimulante au fost desemnate proteine \u200b\u200bG s (stimulează) și proteine \u200b\u200bG inhibitoare (proteine \u200b\u200bG (inhibă) (Fig. 11).
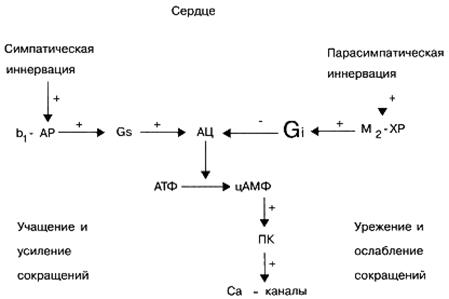
Fig. 11. Mecanismul schimbărilor în frecvența și puterea contracțiilor inimii în timpul stimulării inervației simpatice și parasimpatice.
Toxina coleră activează proteinele C s (aceasta duce la activarea adenilat ciclazei și în holeră se manifestă prin secreția de lichid prin epiteliul intestinal).
Toxina Pertussis activează proteinele G i.
Atunci când receptorii colinergici M 1, receptorii colinergici M 3, α 1 -adrenoreceptorii sunt excitați prin proteinele G q, fosfolipasa C este activată, ceea ce contribuie la formarea inozitol-1,4,5-trifosfat și diacilglicerol din fosfatidilinositol-4,5-difosfat .
Inositolul-1,4,5-trifosfat acționează asupra receptorilor membranei reticulului sarcoplasmic sensibile la aceasta și stimulează eliberarea ionilor de Ca 2+ din reticulul sarcoplasmic (Fig. 12). Prin stimularea αι-adrenoreceptorilor vaselor de sânge, aceasta duce la o contracție a mușchilor netezi ai vaselor și la îngustarea vaselor (Fig. 13).

Fig. 12. Efectul fosfolipazei C asupra nivelului C a2 + citoplasmatic.
Sensibilitatea receptorilor la agoniști și numărul de receptori sunt în continuă schimbare. Deci, după stimularea β 1 \u200b\u200b-adrenoreceptorilor de către un agonist β 1 -adrenoreceptor, fosforilatul de către un receptor kinaza special, se leagă de proteina β-arrestină și în acest complex își pierd capacitatea de a interacționa cu proteinele G (desensibilizarea receptorilor). Complexul de β 1 -adrenoreceptori cu β-arrestină este absorbit de celulă prin endocitoză (interiorizarea receptorilor) și este capturat de endosomi și lizozomi. În endosomi, moleculele β-restin sunt detașate de receptorii care se reintegrează în membrana celulară; sensibilitatea receptorilor la agoniști este restabilită (resensibilizarea receptorilor). În lizozomi, distrugerea moleculelor receptorilor are loc (reglarea în jos) (Fig. 14).

Fig. 13. Mecanismul de contracție a mușchilor netezi ai vaselor de sânge în timpul stimulării inervației simpatice. FLS - fosfolipază C; FIF 2 - fosfatidilinositol-4,5-difosfat; Dacă 3 - inozitol-1,4,5-trifosfat; CP - reticul sarcoplasmic; MLCK - kinazele lanțurilor ușoare ale miozinei.

Fig. 14. Desensibilizarea și reglarea în jos a receptorilor β-adrenergici.
K receptorii intracelulari sunt incluși receptorii corticosteroizi și hormonilor sexuali. În special, receptorii glucocorticoizi sunt localizați în citoplasma celulelor. După ce glucocorticoidul este combinat cu receptorii citoplasmici, complexul glucocorticoid - receptor pătrunde în nucleu și afectează expresia diferitelor gene.
Capacitatea substanțelor de a se lega de receptori (tendința substanțelor de a se lega de receptori) este indicată prin termenul „ afinitate“. În raport cu aceiași receptori, afinitatea diferitelor substanțe poate fi diferită. Pentru a caracteriza afinitatea, utilizați indicatorul pK D este logaritmul negativ al constantei de disociere, adică concentrația substanței la care sunt ocupate 50% dintre receptori.
Activitate internă - capacitatea substanțelor de a stimula receptorii; determinat de efect farmacologicasociate cu activarea receptorilor.
În condiții normale, nu există o corelație directă între afinitate și activitatea internă. O substanță poate ocupa toți receptorii și poate provoca un efect slab, și invers, o substanță poate ocupa 10% din receptori și poate provoca efectul maxim pentru acest sistem.
agoniști - substanțe cu afinitate și activitate internă.
Agoniști complete posedă afinitate și activitate internă maximă (pot provoca efectul maxim pentru un anumit sistem), chiar dacă ocupă o parte din receptorii specifici.
Agoniști parțiali (parțiali) posedă afinitate și mai puțin decât maximă activitate internă (ele pot provoca doar mai puțin decât efecte maxime, chiar dacă ocupă 100% din receptorii specifici).
antagoniștiposedă afinitate, dar nu posedă activitate internă și interferează cu acțiunea agoniștilor deplini sau parțiali (îndepărtează agoniștii de la comunicarea cu receptorii).
Dacă acțiunea antagonistului este eliminată prin creșterea dozei de agonist, un astfel de antagonism este numit competitiv.
Agoniștii parțiali pot fi antagoniști ai agoniștilor deplini. În absența unui agonist complet, un agonist parțial stimulează receptorii și determină un efect slab. Când interacționează cu un agonist complet, un agonist parțial ocupă receptori și inhibă acțiunea unui agonist complet. În acest caz, efectul unui agonist complet este slăbit.
De exemplu, pindololul, un agonist parțial β-adrenoreceptor, provoacă tahicardie slabă în absența efectelor inervației simpatice asupra inimii. Dar, cu creșterea tonului inervației simpatice, pindololul acționează ca un veritabil blocant β și provoacă bradicardie. Acest lucru se datorează faptului că pindololul agonist parțial slăbește efectul norepinefrinei mediator, care este un agonist complet în ceea ce privește receptorii β-adrenergici ai inimii.
Agoniști antagoniști - substanțe care acționează diferit asupra subtipurilor acelorași receptori: stimulează unele subtipuri de receptori și blochează altele. De exemplu, nalbupina analgezică narcotică are efecte diferite asupra subtipurilor de receptori opioizi. Nalbuphine stimulează receptorii κ (și, prin urmare, reduce sensibilitatea la durere) și receptorii μ se blochează (și, prin urmare, mai puțin periculoși în ceea ce privește dependența de droguri).
Un exemplu de efect al substanțelor asupra enzime poate exista o acțiune a agenților anticolinesteraza care blochează acetilcolinesteraza (o enzimă care descompune acetilcolina) și îmbunătățește și întinde acțiunea acetilcolinei.
Substanțe medicinale cunoscute care stimulează sau blochează canale ionice membrane celulare, adică canale care conduc selectiv ioni de Na +, K +, Ca 2+ (sodiu, potasiu, calciu), etc. De exemplu:
Anestezicele locale blochează canalele Na +;
Medicamentele antiaritmice de clasa I (chinidină, lidocaină) blochează canalele Na +;
Minoxidil activează canalele K +;
Agenții hipoglicemici din grupul derivaților de sulfoniluree blochează canalele K + dependente de ATP;
Verapamil, nifedipină blochează canale Ca 2+.
Un exemplu de efect al substanțelor asupra sisteme de transport poate exista o acțiune:
Reserpina (blochează absorbția veziculară a dopaminei și norepinefrinei);
Glicozide cardiace (inhibă Na + / K + -ATPază);
Antidepresive triciclice (blochează absorbția neuronală inversă a norepinefrinei și serotoninei);
Blocante ale pompei de protoni (omeprazol etc.).
Alte mecanisme de acțiune sunt posibile. De exemplu, manitolul diuretic crește diureza prin creșterea presiunii osmotice în tubii renali. Agentul anti-aterosclerotic - colestipolul - leagă (sechestrează) acizii biliari, previne absorbția lor în intestin și, prin urmare, formarea de acizi biliari din colesterolul din ficat este activată și nivelul colesterolului în hepatocite scade.
Mecanismele de acțiune ale diverselor medicamente au fost studiate în diferite grade. În procesul studierii, ideile despre mecanismele de acțiune nu numai că pot deveni mai complicate, dar se pot schimba semnificativ.
Conceptul de „ localizarea acțiunii"Înseamnă locul principal (locul) de acțiune al anumitor substanțe medicinale. De exemplu, glicozidele cardiace acționează în principal asupra inimii.
La conceptul de „ tipuri de acțiune»În legătură cu acțiunile locale și generale (resorptive), acțiune reflexă, acțiune primară și secundară, acțiune directă și indirectă.
Un exemplu de acțiune locală poate fi acțiunea anestezicelor locale.
Majoritatea medicamentelor au un efect general (resorptiv), care se dezvoltă de obicei după absorbția (resorbția) unei substanțe în sânge și distribuirea acesteia în organism.
Atât cu acțiuni locale, cât și cu acțiuni resorctive, substanțele pot excita diverși receptori sensibili și pot provoca reacții reflexe.
Efectul principal al unei substanțe medicinale este efectele sale, care sunt utilizate în fiecare caz. Toate celelalte efecte sunt evaluate ca manifestări ale reacțiilor adverse.
Substanțele medicinale se pot exercita asupra anumitor organe acțiune directă. În plus, efectul medicamentelor poate fi indirect. De exemplu, glicozidele cardiace au un efect direct asupra inimii, dar, îmbunătățind funcționarea inimii, aceste substanțe cresc aportul de sânge și funcțiile altor organe (efect indirect).