Didžioji dauguma vaistų turi gydomąjį poveikį keičiant ląstelių, kurios evoliucijos metu buvo sukurtos kūne, fiziologinių sistemų aktyvumą. Paprastai veikiant vaistinei medžiagai organizme, naujo tipo ląstelės neatsiranda, keičiasi tik įvairių natūralių procesų greitis. Fiziologinių procesų slopinimas ar sužadinimas lemia atitinkamų kūno audinių funkcijų sumažėjimą ar sustiprėjimą.
Vaistai gali veikti specifinius receptorius, fermentus, ląstelių membranas arba tiesiogiai sąveikauti su ląstelių medžiagomis. Narkotikų veikimo mechanizmai išsamiai tiriami bendrosios ar eksperimentinės farmakologijos metu. Žemiau pateikiame tik keletą pagrindinių narkotikų veikimo mechanizmų pavyzdžių.
Poveikis specifiniams receptoriams. Receptoriai yra makromolekulinės struktūros, selektyviai jautrios tam tikriems cheminiams junginiams. Cheminių medžiagų sąveika su receptoriais lemia biocheminių ir fiziologinių pokyčių atsiradimą organizme, kurie pasireiškia tam tikru klinikiniu poveikiu.
Vaistai, kurie tiesiogiai sužadina ar padidina funkcinį receptorių aktyvumą, vadinami agonistais, o medžiagos, slopinančios specifinių agonistų veikimą, vadinamos antagonistais. Antagonizmas gali būti konkurencingas ir nekonkurencingas. Pirmuoju atveju vaistinė medžiaga konkuruoja su natūraliu reguliatoriumi (tarpininku) dėl jungimosi vietų specifiniuose receptoriuose. Konkurencinio antagonisto sukeltą receptorių blokadą galima pašalinti didelėmis agonisto ar natūralaus tarpininko dozėmis.
Receptorių įvairovė yra padalinta pagal jautrumą natūraliems mediatoriams ir jų antagonistams. Pavyzdžiui, acetilcholinui jautrūs receptoriai yra vadinami cholinerginiais, adrenalinui jautrūs receptoriai yra vadinami adrenerginiais. Kalbant apie jautrumą muskarinui ir nikotinui, cholinerginiai receptoriai yra suskirstyti į jautrius, jautrius muskarinams (m-cholinerginius receptorius) ir nikotinui jautrius (n-cholinerginius receptorius). H-cholinerginiai receptoriai yra nevienalyčiai. Yra nustatyta, kad jų skirtumas yra jautrumas įvairioms medžiagoms. Yra n-cholinerginiai receptoriai, esantys autonominės nervų sistemos ganglijose, ir n-cholinerginiai receptoriai, susiaurėjusio raumens. Yra žinomi įvairūs adrenerginių receptorių potipiai, žymimi graikiškomis raidėmis α1, α 2, β1, β2.
Taip pat išskirti H1 ir H2 histaminai, dopaminas, serotoninas, opioidai ir kiti receptoriai.
Poveikis fermentų aktyvumui. Kai kurie vaistai padidina arba slopina specifinių fermentų aktyvumą. Pavyzdžiui, fizostigminas ir neostigminas mažina cholinesterazės, kuri naikina acetilcholiną, aktyvumą ir suteikia poveikį, kuris būdingas parasimpatinės nervų sistemos sužadinimui. Monoaminooksidazės inhibitoriai (iprazidas, nialamidas), neleidžiantys sunaikinti adrenalino, padidina simpatinės nervų sistemos veiklą. Fenobarbitalis ir ziksorinas, padidindami kepenų gliukuronilo transferazės aktyvumą, sumažina bilirubino kiekį kraujyje.
Fizikinis ir cheminis poveikis ląstelių membranoms. Nervų ir raumenų sistemų ląstelių aktyvumas priklauso nuo jonų, lemiančių transmembraninį elektrinį potencialą, srauto. Kai kurie vaistai keičia jonų transportą.
Taigi antiaritminiai, prieštraukuliniai vaistai, vaistai bendrajai anestezijai.
Tiesioginė cheminė sąveika. Vaistai gali tiesiogiai sąveikauti su mažomis molekulėmis ar jonais ląstelių viduje. Pavyzdžiui, etilendiamintetraacto rūgštis (EDTA) stipriai jungiasi su švino jonais. Tiesioginės cheminės sąveikos principas grindžiamas daugelio priešnuodžių naudojimu apsinuodijus cheminėmis medžiagomis. Kitas pavyzdys - druskos rūgšties neutralizavimas antacidais.
Farmakodinamika
Jis tiria vaistų veikimo mechanizmą, taip pat jų biocheminį ir fiziologinį poveikį. Jos užduotys apima cheminės ir fizinės sąveikos tarp vaisto ir tikslinės ląstelės aprašymą, taip pat visą jo farmakologinio poveikio spektrą ir sunkumą. Farmakodinaminių modelių žinojimas leidžia pasirinkti tinkamus vaistus. Farmakodinaminiai tyrimai suteikia gilesnį supratimą apie biocheminių ir fiziologinių procesų reguliavimą organizme (Katzung B.G., 1998; Lawrence D.R. ir kt., 2002).
Daugelio vaistų veikimą skatina jų prisijungimas prie organizmo makromolekulių. Šių makromolekulių funkcinės būklės pasikeitimas savo ruožtu suaktyvina biocheminių ir fiziologinių reakcijų grandinę, kuri virsta farmakologiniu poveikiu. Makromolekulės, su kuriomis sąveikauja cheminės medžiagos, vadinamos receptoriais. Taigi bet kurios funkciškai aktyvios makromolekulės gali tarnauti kaip vaistų receptoriai. Iš šio teiginio išplaukia kelios svarbios pasekmės. Pirmiausia, pasitelkę vaistus, galite pakeisti bet kurio fiziologinio proceso organizme greitį. Antra, vaistai keičia tik natūralias ląstelės fiziologines funkcijas, nesuteikdami jai naujų savybių.
Receptoriai
Dauguma receptorių yra baltymai. Tai yra hormonų receptoriai, augimo faktoriai, mediatoriai, baltymai, dalyvaujantys svarbiausiose medžiagų apykaitos ir reguliavimo reakcijose (dihidrofolato reduktazė, acetilcholinesterazė), transportiniai baltymai (Na +, K + -ATPazė), struktūriniai baltymai (tubulinas). Ląstelės komponentai, kurie yra skirtingo cheminio pobūdžio, pavyzdžiui, nukleino rūgštys, su kuriomis sąveikauja priešnavikiniai agentai, taip pat gali veikti kaip receptoriai.
Endogeninių reguliavimo veiksnių receptoriai - hormonai, mediatoriai ir kt. - turi farmakologinę reikšmę. Šie receptoriai yra daugelio vaistų taikiniai, paprastai veikiantys selektyviai dėl didelio endogeninių ligandų receptorių specifiškumo. Vaistai, kurie, prisijungdami prie receptorių, atkuria endogeninio ligando fiziologinį poveikį, vadinami aganistais arba stimuliatoriais. Vaistai, kurie nesukelia šio efekto, bet slopina endogeninių ligandų surišimą, vadinami antagonistais arba blokatoriais. Medžiagos, kurių poveikis yra mažiau ryškus nei agonistų poveikis, vadinamos daliniais agonistais. Preparatai, stabilizuojantys receptorių neaktyvia forma, yra klasifikuojami kaip atvirkštiniai agonistai.
Struktūrinė ir funkcinė priklausomybė
Cheminė vaisto struktūra gana griežtai nustato jo afinitetą receptoriams ir vidinį aktyvumą. Nedidelis cheminės struktūros pasikeitimas gali reikšmingai paveikti farmakologines savybes.
Naujų vaistų sintezė daugiausia paremta tuo. Kadangi cheminis modifikavimas nebūtinai vienodai veikia visas farmakologines savybes, galima pagerinti vaisto veiksmingumą ir saugumą, padidinti jo selektyvumą ir pagerinti farmakokinetines savybes. Pavyzdžiui, daugelis klinikoje naudojamų hormonų ir mediatorių antagonistų yra sintetinami chemiškai modifikuojant endogenines medžiagas.
Vaistų vartojimo taškai
Kadangi vaistų poveikį tarpininkauja receptoriai, vaisto vartojimo tašką lemia ne tik jo pasiskirstymo ypatybės, bet ir receptorių lokalizacija, o farmakologinis poveikis priklauso nuo šių receptorių funkcinės reikšmės. Vaistų, kurių receptoriai yra paplitę daugelyje organų ir audinių, farmakologinis poveikis yra įvairus. Jei šie receptoriai atlieka ląstelėms gyvybiškai svarbią funkciją, tai ne tik sunku vartoti vaistą terapiniais tikslais, bet ir nesaugu. Nepaisant to, tokie vaistai gali turėti didelę klinikinę reikšmę. Taigi širdies glikozidai, plačiai naudojami sergant širdies nepakankamumu, keičia jonų pernešimą per ląstelės membraną, nuo kurios priklauso gyvybinis ląstelės aktyvumas. Jie pasižymi siauru terapiniu diapazonu ir yra labai toksiški. Kitas pavyzdys yra priešnavikiniai vaistai. Jei receptoriai, su kuriais sąveikauja vaistas, yra tik kelių rūšių diferencijuotose ląstelėse, jo poveikis yra selektyvesnis. Šie vaistai gali sukelti mažiau nepageidaujamų reakcijų, tačiau vis dėlto šie vaistai gali būti toksiški, jei jų receptoriai atlieka gyvybiškai svarbią funkciją. Kai kurie biologiniai nuodai (botulino toksinas ir kt.) Veikia panašiai. Be to, net jei tiesioginis farmakologinis poveikis yra selektyvus, jo pasekmės gali būti įvairesnės.
Endogeninių reguliavimo faktorių receptoriai
Terminas receptorius reiškia bet kokį makromolekulinį ląstelės komponentą, su kuriuo jungiasi vaistas. Vienas iš svarbiausių vaistų receptorių yra ląstelių baltymai, kurie tarnauja kaip endogeninių reguliavimo veiksnių receptoriai - hormonai, augimo faktoriai, mediatoriai. Jungdamiesi prie endogeninio ligando, receptoriai perduoda signalą iš jo į tikslinę ląstelę.
Iš receptoriaus signalas patenka į ląstelių taikinius (efektoriniai baltymai) tiesiogiai arba per tarpines signalines molekules - baltymų konverterius. Receptorius, baltymų konverteriai ir efektoriniai baltymai sudaro receptoriaus-efektoriaus sistemą. Artimiausias efektoriaus baltymas signalo perdavimo grandinėje dažnai yra ne galinis efektorius (tiesiogiai veikiantis ląstelių funkcijas), bet fermentas arba transportinis baltymas, dalyvaujantis formuojant, pernešant ar inaktyvuojant antrąjį tarpininką - joną ar mažą molekulę. Antrasis tarpininkas savo ruožtu perduoda informaciją į įvairius tarpląstelinius taikinius, užtikrindamas, kad jie tuo pačiu metu reaguoja į vieno receptoriaus signalą.
Receptoriai, konvertuojantys baltymus, ir efektoriniai baltymai ne tik perduoda informaciją. Jie taip pat koordinuoja signalus iš skirtingų ligandų, viena vertus, ir visus šiuos signalus, susijusius su medžiagų apykaitos procesais ląstelėje.
Veikdami kaip katalizatoriai, receptoriai sustiprina biologinį signalą. Dėl šios svarbios savybės jie yra puikūs vaistų tikslai. Tačiau signalo stiprintuvai yra ne tik receptoriai, turintys fermentinį aktyvumą, bet ir visi žinomi receptoriai. Iš tiesų, kai viena ligando molekulė jungiasi prie receptoriaus, konjuguoto su jonų kanalu, daugelis jonų praeina pro pastarąjį. Tas pats pasakytina ir apie steroidinių hormonų receptorius: viena hormono molekulė sukelia daugelio mRNR kopijų, kurių pagrindu susintetinta daugybė baltymų molekulių, transkripciją.
Atsižvelgiant į struktūrą ir veikimo mechanizmą, biologiškai aktyvių medžiagų receptoriai yra suskirstyti į keletą klasių. Šių užsiėmimų skaičius yra nedidelis.
Fermentiniai receptoriai
Didžiausia receptorių grupė, turinti fermentinį aktyvumą, yra membraniniai receptoriai, turintys savo baltymų kinazės aktyvumą. Jie fosforilina įvairius efektorinius baltymus, esančius ląstelės membranos viduje. Dėl to keičiasi šių baltymų funkcija arba jų sąveika su kitais baltymais.
Yra dar viena receptorių, turinčių baltymų kinazės aktyvumą, klasė - tai receptoriai, konjuguoti su baltymų kinazėmis. Jiems trūksta tarpląstelinio katalitinio domeno, tačiau, sąveikaudami su agonistais, jie jungiasi arba aktyvuoja tarpląstelines baltymų kinazes vidiniame membranos paviršiuje. Tai yra neurotrofinių veiksnių receptoriai ir T ir B limfocitų, susidedančių iš kelių subvienetų, antigenus atpažįstantys receptoriai. Pastarieji taip pat sąveikauja su fosfotirozino fosfatais. Kitų receptorių, neturinčių intraląstelinio efektoriaus domeno, funkciją gali tarpininkauti kai kurie kiti efektoriniai baltymai.
Kiti receptoriai, turintys savo fermentinį aktyvumą, turi panašią struktūrą. Tai apima, pavyzdžiui, receptorius, turinčius savo fosfotirozino fosfatazės aktyvumą: jų tarpląstelinis domenas aminorūgščių seka yra panašus į adhezijos molekules. Daugeliui receptorių, turinčių savo fosfotyrosine fosfatazės aktyvumą, endogeniniai ligadai nežinomi. Tačiau, remiantis genetiniais ir biocheminiais tyrimais, atliktais su skirtingų tipų ląstelėmis, svarbų vaidmenį vaidina fermentinis šių receptorių aktyvumas. Prieširdžių natriuretinio hormono receptorių, kitų NUP, taip pat guanilino receptorių tarpląstelinis domenas turi savo guanilato ciklazės aktyvumą ir sintezuoja cGMP, kuris veikia kaip antrasis tarpininkas. Galbūt yra ir kitų receptorių, turinčių savo fermentinį aktyvumą.
Jonų kanalais sujungti receptoriai
Kai kurių mediatorių receptoriai yra tiesiogiai susiję su jonų kanalais, sąveikaudami su ligandu, selektyviai praeina tam tikrus jonus per ląstelės membraną (chemiškai jautrūs kanalai, jonotropiniai receptorių kanalai, jonotropiniai receptoriai).
G baltymų jungiami receptoriai
Tai gana didelė receptorių klasė, kuri sąveikauja su efektoriais per G baltymus (baltymai, kuriuose guanino difosfatas (GDF) pakeičia guanino trifosfatą (GTP). Tai apima daugelio biogeninių aminų, lipidus signalizuojančių molekulių (ypač eikosanoidų) ir įvairių peptidų receptorius. fermentai (adenilato ciklazė, fosfolipazė C) ir kalio bei kalcio membranos kanalai veikia kaip efektoriai. Didelis skaičius ir svarbus fiziologinis receptorių, sujungtų su G baltymais, vaidmuo daro juos puikius. mano tikslai dėl narkotikų: maždaug pusė visų gydytojų išrašytų vaistų (išskyrus antibiotikus) veikia šiuos receptorius.
Ląstelė ant savo paviršiaus gali nešti iki 20 receptorių, kurių kiekvienas selektyviai sąveikauja su vienu ar daugiau G-baltymų tipų (jie skiriasi skirtingais α-subvienetų tipais). Α-subvienetas gali sąveikauti su vienu ar daugiau efektorinių baltymų, o tai leidžia koordinuoti signalus iš skirtingų ligandų receptorių, naudojant vieną G-baltymą. Kita vertus, vienas receptorius gali suaktyvinti kelis tarpląstelinio signalo perdavimo mechanizmus, suaktyvindamas kelių tipų G baltymus ir veikdamas skirtingus efektorinius baltymus per tą patį α-subvienetą. Tokia sudėtinga signalų divergencijos ir konvergencijos sistema suteikia galimybę lanksčiai reguliuoti ląstelių funkcijas (Ross, 1992).
Tarpląsteliniai receptoriai
Steroidinių ir skydliaukės hormonų, kalcitriolio ir retinoidų receptoriai yra tirpūs tarpląsteliniai DNR jungiantys baltymai, kurie reguliuoja tam tikrų genų transkripciją (Mangelsdorf ir kt., 1994). Šie receptoriai priklauso ligandui jautrių transkripcijos reguliatorių šeimai. Transkripcijos veiksnių funkciją reguliuoja fosforilinimas, sąveika su ląstelės baltymais, metabolitais ir kitais ląstelės reguliavimo elementais.
Antrosios tarpinės sistemos
cAMP. Antrinės tarpinės sistemos taip pat dalyvauja integruojant išorinius signalus. Nors yra daug daugiau žinomų receptorių ir baltymus signalizuojančių molekulių nei antrieji mediatoriai, pastarieji dalyvauja daugelyje ląstelių signalo perdavimo būdų. Labiausiai ištirti antrieji tarpininkai yra cAMP, cGMP, Ca 2+, IF 3 (inozitolio trifosfatas), DAG (diacilglicerolis), NO. Ši heterogeninių junginių grupė nuolat auga. Antrieji tarpininkai sąveikauja tiesiogiai (keisdami vienas kito metabolizmą) arba netiesiogiai (veikdami tuos pačius tarpląstelinius taikinius). Antrųjų tarpininkų funkcija, taip pat jų formavimo (ar išsiskyrimo), skaidymo ir išskyrimo iš ląstelės reguliavimas, yra patogiai nagrinėjama cAMP pavyzdžiu. Šis antrasis tarpininkas yra sintetinamas veikiant adenilato citazei, suaktyvinus daugelį receptorių, konjuguotų su G baltymais. G s baltymas aktyvina adenilato ciklazę, G i baltymas slopina.
Yra mažiausiai 10 audiniui būdingų adenilato ciklotazės izoformų, kurios skiriasi aktyvumo reguliavimo mechanizmais.
Paprastai cAMP aktyvina baltymų kinazes A (nuo cAMP priklausomas baltymų kinazes), nedidelę susijusių baltymų grupę. Šios baltymų kinazės savo ruožtu fosforilina ne tik galutinius tarpląstelinius taikinius (fermentus, transportinius baltymus), bet ir kitas baltymų kinazes bei kitus reguliuojančius baltymus. Pastarieji apima, pavyzdžiui, transkripcijos veiksnius. Jie yra atsakingi už cAMP tarpininkaujamą genų transkripcijos reguliavimą, užtikrinant uždelstą ląstelės atsaką į signalą. Be baltymų kinazių aktyvavimo, cAMP veikia tiesiogiai katijoninės membranos kanalus, kurie vaidina svarbų vaidmenį, visų pirma, veikiant neuronams. Taigi signalas iš cAMP sukelia biocheminių pokyčių grandinę tikslinėje ląstelėje.
Kalcis Kitas gerai ištirtas antrasis tarpininkas yra tarpląstelinis Ca 2+. Ca 2+ jonai į citoplazmą patenka įvairiais būdais: per membraninius kanalus (priklausomus nuo G baltymų, nuo įtampos, reguliuojamus K + arba Ca 2+ sim), taip pat per kanalus, esančius konkrečiose endoplazminio retikulumo vietose ir atsiveriančius veikiant. IF 3 ir skeleto raumenyse dėl membranos depoliarizacijos. Kalcis iš citozolinės plazmos pašalinamas dviem būdais: jis absorbuojamas endoplazminio retikulumo būdu arba pašalinamas iš ląstelės. Ca 2+ perduoda signalus daug didesniam skaičiui baltymų nei cAMP - fermentai, dalyvaujantys ląstelių metabolizme, baltymų kinazės, kalcį jungiantys baltymai. Pastarosios sąveikauja su kitais galutiniais ir tarpiniais efektoriais.
Receptorių reguliavimas
Receptoriai ne tik kontroliuoja fiziologines ir biochemines funkcijas, bet ir tarnauja kaip reguliavimo objektai. Šis reguliavimas atliekamas jų makromolekulių sintezės ir skilimo lygmeniu, formuojant kovalentinius ryšius su kitomis molekulėmis, sąveikaujant su reguliuojančiais baltymais ir receptorių judėjimu. Konvertuojantys baltymai ir efektoriniai baltymai taip pat yra reguliuojami. Normatyviniai signalai gali būti gaunami iš tarpląstelinio perdavimo būdų, aktyvuojamų stimuliuojant patį receptorių (per grįžtamojo ryšio mechanizmą), taip pat iš kitų receptorių (tiesiogiai ar netiesiogiai).
Ilgalaikis vaistų receptorių stimuliavimas paprastai lemia reakcijos į jį sumažėjimą - esant tokiai pačiai koncentracijai, vaistas sukelia mažiau ryškų poveikį. Šis reiškinys, vadinamas desensibilizacija, refrakcija ir priklausomybe, vaidina svarbų vaidmenį klinikinėje praktikoje: pavyzdžiui, ilgai gydant β-adrenerginius agonistus pacientams, sergantiems AD, sumažėja reakcijos į šiuos vaistus sunkumas.
Homologinis desensibilizacija taikoma tik stimuliuotiems receptoriams ir būdinga ligandui. Dėl heterologinio desensibilizacijos sumažėja reakcijos į kitus ligandus, kurių receptoriai veikia tuo pačiu tarpląsteliniu signalo perdavimo keliu, sunkumas. Pirmuoju atveju neigiamą grįžtamąjį ryšį teikia poveikis pačiam receptoriui (fosforilinimas, proteolizė, sumažėjusi sintezė), antruoju atveju, be receptoriaus, jis gali paveikti ir kitus baltymus, susijusius su tarpląstelinio signalo perdavimu.
Priešingai, jei receptoriai ilgą laiką nėra stimuliuojami, padidėja jų jautrumas agonistams (pavyzdžiui, ilgą laiką gydant β-adrenoblokatoriaus propronololiu, padidėja β-adrenoblokatorių jautrumas β-adrenostimuliatoriams).
Sutrikimai dėl sutrikusios receptorių funkcijos
Be individualių jautrumo vaistams skirtumų, yra ir ligų, kurias sukelia kai kurių ląstelių signalo perdavimo iš receptorių į efektorių mechanizmo tam tikrų komponentų disfunkcija. Praradus labai specializuotų receptorių funkciją, fenotipinės ligos apraiškos gali būti ribotos (pavyzdžiui, sėklidžių feminizacija susijusi su genetiniu androgenų receptorių nebuvimu ar struktūriniais defektais). Jei pažeidžiamas universalesnis ląstelių signalo perdavimo mechanizmas, ligos simptomai būna įvairesni, pavyzdžiui, sergant myasthenia gravis ir kai kuriomis insulinui atsparaus cukrinio diabeto formomis, kurias atitinkamai sukelia N-cholinerginių receptorių ir insulino receptorių autoimuniniai disfunkcijos. Bet kurio komponento, susijusio su signalo perdavimu iš daugelio receptorių, defektai sukelia daugybinius endokrininius sutrikimus. Pavyzdys yra heterozigotinė G baltymų, aktyvinančių adenilato ciklazę, trūkumo forma visose ląstelėse (Spiegel ir Weinstein, 1995). Tikėtina, kad homozigotinė šio baltymo trūkumo forma gali baigtis mirtimi.
Receptorių struktūros ar lokalizacijos sutrikimai gali pasireikšti kaip susilpnėjusi ar sustiprėjusi reakcija į vaistą, taip pat kaip kitas nepageidaujamas poveikis.
Mutacijos, koduojančios genų receptorius, gali pakeisti tiek reakciją į vienkartinį vaisto vartojimą, tiek ilgalaikio gydymo efektyvumą. Pavyzdžiui, β-adrenerginių receptorių, atsakingų už bronchų lygiųjų raumenų atpalaidavimą ir kvėpavimo takų atsparumo reguliavimą, trūkumas padidina šių receptorių jautrumo β-adrenostimuliatoriams sumažėjimą ilgalaikio AD sergančių pacientų gydymo metu. Nustačius mutacijas, sukeliančias sutrikusią receptorių funkciją, ir klonuojant atitinkamus genus, bus įmanoma sukurti tokių ligų gydymo metodus.
Receptorių klasifikacija
Tradiciškai vaistų receptoriai buvo nustatomi ir klasifikuojami remiantis selektyvių agonistų (stimuliatorių) ir antagonistų (blokatorių), kurie veikia šiuos receptorius, poveikiu ir santykiniu aktyvumu. Pavyzdžiui, acetilcholino poveikis, kuris atkuriamas sąveikaujant su cholinerginiais muskarino alkaloido receptoriais ir kurį blokuoja atropinas, yra vadinamas muskarininiu poveikiu, o poveikis, kuris atkuriamas sąveikaujant su cholinerginiais nikotino receptoriais, vadinamas nikotininiu poveikiu. Receptoriai, tarpininkaujantys muskarino ir nikotino poveikiui, vadinami atitinkamai M ir N cholinerginiais receptoriais. Nors tokia klasifikacija paprastai neatspindi vaistų veikimo mechanizmo, patogu susisteminti jų poveikį. Iš tiesų teiginys, kad vaistas stimuliuoja tam tikro tipo receptorius, tuo pačiu lemia šio narkotiko ir medžiagų, kurios sustiprina arba silpnina šį poveikį, spektrą. Tačiau tokių teiginių galiojimas gali keistis nustačius naujų tipų ir potipių receptorius, atradus papildomus vaistų veikimo mechanizmus ar anksčiau nežinomą šalutinį poveikį.
Receptorių potipiai
Atsiradus vis gausėjančiai labai selektyvių vaistų įvairovei, tapo akivaizdu, kad anksčiau žinomi receptorių tipai yra suskirstyti į daugelį potipių. Molekulinio klonavimo metodai tapo reikšminga pagalba tiriant naujus receptorių potipius, o rekombinantinių receptorių paruošimas palengvino vaistų, selektyviai veikiančių šiuos receptorius, kūrimą. Skirtingi, bet susiję receptorių potipiai dažnai (nors ne visada) sąveikauja su skirtingais agonistais ir antagonistais. Receptoriai, kuriems nenustatyta selektyviųjų agonistų ar antagonistų, paprastai priklauso ne vienam potipiui, o to paties receptoriaus izoformoms.Skiriami potipiai taip pat gali skirtis tarpląstelinio signalo perdavimo mechanizmuose. M 1 ir M 3 cholinerginiai receptoriai, pavyzdžiui, veikia per baltymą G q, kuris aktyvina fosfolipazę C, netiesiogiai sukelia Ca 2+ išsiskyrimą iš viduląstelinių saugyklų, o M 2 ir M 4 cholinerginius receptorius per baltymą G i, kuris slopina adenilato ciklazę. Tuo pačiu metu receptorių suskirstymą į tipus ir potipius dažnai lemia ne veikimo mechanizmas, o atsitiktinis pasirinkimas arba jis grindžiamas nusistovėjusiomis idėjomis. Taigi α 1 -, α 2 - ir β-adrenerginiai receptoriai skiriasi pagal reakciją į vaistus ir signalų perdavimą (atitinkamai aktyvina baltymus G i, G q ir G s), nors α ir β adrenerginiai receptoriai yra skirtingų tipų, ir α1 - ir α 2 -adrenoreceptoriai - skirtingiems to paties tipo potipiams. Α 1 -adrenoreceptorių α 1A, α 1B ir α 1D izoformos mažai skiriasi savo biocheminėmis savybėmis; tas pats būdinga β-adrenerginių receptorių izoformrazių potipiams (β 1, β 2 ir β 3).
Skirtumai tarp receptorių potipių naudojami kuriant labai selektyvius vaistus, pavyzdžiui, vaistus, turinčius skirtingą poveikį tam pačiam audiniui dėl prisijungimo prie receptorių potipių, kurie skiriasi tarpląstelinio signalo perdavimo mechanizmais. Be to, vaistai gali selektyviai nukreipti tam tikras ląsteles ar audinius, išreiškiančius potipio receptorius. Kuo didesnis vaistų selektyvumas (atsižvelgiant į tam tikrą audinį ar į tam tikrą poveikį), tuo palankesnis yra jo naudos ir nepageidaujamo poveikio santykis.
Naudojant molekulinius genetinius metodus, buvo atrastos ne tik skirtingos receptorių izoformos, bet ir genai, koduojantys naujus, anksčiau nežinomus receptorius. Daugelis šių receptorių jau priskiriami vienai ar kitai žinomai klasei, o jų funkcija buvo tiriama naudojant atitinkamus ligandus. Tačiau kai kuriems receptoriams dar nebuvo rasta ligandų.
Daugelio to paties receptoriaus izoformų, užkoduotų skirtingų genų, atradimas (ypač jei izoformos nesiskiria tarpląstelinio signalo perdavimo mechanizmuose ir sąveikauja su tais pačiais endogeniniais ligandais) leidžia receptorių ekspresiją skirtingose \u200b\u200bląstelėse nepriklausomai reguliuoti atsižvelgiant į kūno poreikius skirtinguose. amžiaus laikotarpiai.
Nereceptinių vaistų veikimas
Ne visi vaistai veikia per makromolekulines struktūras - receptorius. Kai kurie vaistai sąveikauja su mažomis molekulėmis ar jonais, esančiais kūne paprastai arba esant vienokiai ar kitokiai patologinei būklei. Taigi, antacidai neutralizuoja druskos rūgštį skrandyje. Mesna (vaistas, greitai pašalinamas pro inkstus ir neutralizuoja laisvuosius radikalus) jungiasi su kai kurių priešnavikinių vaistų aktyviaisiais metabolitais, mažindamas nepageidaujamų reakcijų iš šlapimo takus sunkumą. Daugybė biologiškai neaktyvių medžiagų (pavyzdžiui, manitolio) gali būti įnešta tokiu kiekiu, kad padidėtų biologinių skysčių osmolariumas ir taip būtų pakeistas tarpląstelinių ir tarpląstelinių skysčių pasiskirstymas. Su šių medžiagų pagalba galima padidinti diurezę, padidinti kraujospūdį, pašalinti smegenų edemą. Be to, jie naudojami kaip vidurius laisvinantys vaistai.
Kai kurie vaistai gali integruotis į ląstelės komponentus ir pakeisti jų funkcijas dėl struktūrinių panašumų su medžiagomis, kurios sudaro šiuos komponentus. Pavyzdžiui, purinų ir pirimidinų analogai įterpiami į nukleorūgštis ir naudojami kaip antivirusiniai ir priešnavikiniai agentai.
A.P. Viktoras "Klinikinė farmakologija"
Vaistų veikimo mechanizmo pagrindas, kaip taisyklė, yra jų gebėjimas inicijuoti (sukelti) sudėtingus biocheminius n / arba biofizinius procesus, kurie galiausiai keičia ir (arba) optimizuoja tikslinės ląstelės funkcinį aktyvumą.
Vaistai savo poveikį organams ir (arba) tikslinėms ląstelėms gali atlikti:
Tiesioginė cheminė sąveika;
Fizikinė ir cheminė sąveika su ląstelės membrana;
Veiksmai specializuotiems fermentams;
Veiksmai reguliuojančių genų atžvilgiu;
Veiksmai specifiniams receptoriams.
Tiesioginė cheminė sąveika LS. Šis vaistų veikimo mechanizmas yra gana retas ir gali būti realizuotas už ląstelės ribų, pavyzdžiui, skrandžio ar žarnų spindyje. Jos esmė slypi tame, kad vaistai pradeda tiesioginę cheminę reakciją su molekulėmis ir (arba) jonais, kurie organizme susidaro normalioje būsenoje, kai atsiranda patologinė būklė. Tiesioginės cheminės sąveikos pavyzdys yra skrandžio druskos rūgšties neutralizavimo cheminė reakcija vartojant antacidinius vaistus (žr. T. 2, p. 112).
Fizikinė ir cheminė vaistų sąveika su ląstelės membrana. Viena iš pagrindinių citoplazminės membranos funkcijų yra jonų mainų tarp citoplazmos ir tarpląstelinės aplinkos įgyvendinimas. Transmembraniniai jonų mainai taip pat gali vykti per specialius nuo įtampos priklausomus transmembraninius jonų kanalus - natrio, kalio, kalcio, chloro ir kt. Kai kurie vaistai, pasiekdami ląstelės membraną, sąveikauja su šiais kanalais ir keičia jų funkcinį aktyvumą. Taigi, pavyzdžiui, antiaritminis IA klasės vaisto chinidino poveikis pagrįstas jo gebėjimu blokuoti Na + jonų praėjimą per transmembraninius natrio kanalus (žr. T. 2, p. 35).
Vaistų poveikis specializuotiems fermentams. Palyginti nedidelis kiekis vaistų realizuoja savo farmakologinį poveikį keičiant kai kurių specializuotų ląstelių fermentų aktyvumą. Vaistai, didinantys ląstelių fermentų aktyvumą, vadinami fermentų induktoriais. Tokį poveikį turi, pavyzdžiui, migdomosios tabletės ir prieštraukulinis vaistas fenobarbitalis, kuris žymiai padidina mikrosominių kepenų fermentų aktyvumą. Šio fenobarbitalio ir artimo jo LS biologinė reikšmė bus nagrinėjama toliau.
Vaistai, slopinantys specializuotų fermentų veiklą, vadinami fermentų inhibitoriais. Taigi, pavyzdžiui, antidepresantas iš monoaminooksidazės inhibitorių (MAO) grupės, vaistas pirlindolis realizuoja savo antidepresantą, slopindamas MAO fermento aktyvumą centrinėje nervų sistemoje (žr. T. 1, p. 294).
Gebėjimas slopinti fermento acetilcholinesterazės aktyvumą yra anticholinesterazės vaistų, pavyzdžiui, fizostigmino, farmakologinio aktyvumo pagrindas. Yra žinoma, kad fiziologinėmis sąlygomis acetilcholinesterazė inaktyvuoja (sunaikina) acetilcholiną - neuromediatorių, perduodantį sužadinimą parasimpatinės nervų sistemos sinapsėse. Fizostigminas, slopindamas acetilcholinesterazės aktyvumą, skatina kaupimąsi neurotransmiterio acetilcholino parasimpatinės sistemos sinapsėse, dėl to padidėja parasimpatinės nervų sistemos tonusas, pasireiškiantis sisteminiu lygmeniu vystantis bradikardijai, didėjant virškinimo traktui, mažinant kraujo spaudimą (BP). mokinys ir kt.
Vaistai gali grįžtamai ir negrįžtamai sąveikauti su fermentais. Pvz., Vaistas enalaprilis grįžtamai slopina angiotenziną konvertuojančio fermento aktyvumą, kuris visų pirma reiškia kraujospūdžio sumažėjimą, o toksiškos fosforo organinės medžiagos negrįžtamai slopina acetilcholinesterazės aktyvumą.
Vaistų poveikis norminiams genams. Šiuo metu mokslininkai bando sukurti vaistus, kurie realizuotų savo farmakologinį poveikį tiesiogiai veikdami reguliuojančių genų fiziologinį aktyvumą. Ši tendencija atrodo ypač viltinga po to, kai 2000 m. Buvo iššifruota žmogaus genomo struktūra. Manoma, kad selektyvus reguliavimo genų funkcijos normalizavimas veikiant narkotikams leis pasiekti sėkmės gydant daugelį, įskaitant anksčiau nepagydomas, ligas.
Vaistų poveikis receptoriams. Prieš pereinant prie narkotikų sąveikos su receptoriais specifikos, būtina išsiaiškinti, ką turime omenyje terminą „receptorius“ (iš lotyniško recipio - imk, imk).
Nuo fiziologijos eigos žinoma, kad terminas „receptorius“ reiškia labai specializuotas formacijas, gebančias suvokti, transformuoti ir perduoti išorinio signalo energiją nervų sistemai. Tokie receptoriai vadinami sensoriniais (nuo lat. Sensus - pojūtis, pojūtis, suvokimas).
Jutimo receptoriai apima klausos, regos, uoslės, skonio, lytėjimo organų receptorius. Šių organų jutimo receptoriai priklauso vadinamiesiems eksteroreceptoriams.
Jei jutimo organų, reaguojančių į išorinius dirginimo dirgiklius, buvimas buvo žinomas nuo senų senovės, tada jutiminių receptorių buvimas kūne buvo abejotinas iki XIX amžiaus vidurio. Pirmą kartą tokių receptorių buvimą kūne pasiūlė rusų fiziologas I.F.Pionas, kuris triušio eksperimente parodė aortos dirginimo metu kraujospūdžio sumažėjimą 1866 m. Šis atradimas paskatino ieškoti ir tirti receptorius, esančius kūno viduje, ir patys šie receptoriai buvo vadinami interoreceptoriais.
Iki XX amžiaus pradžios buvo atskleistas pakankamas jutiminių interoreceptorių skaičius ir įrodytas jų svarbus vaidmuo reguliuojant fiziologines kūno funkcijas.
1905 m. J. Langley įrodė, kad vaistui tepant ant ląstelės membranos, atsiranda farmakologinis poveikis, jei jis taikomas tik tam tikroje jo vietoje. Be to, ši svetainė sudaro tik nedidelę viso ląstelės paviršiaus ploto dalį. Šis pastebėjimas leido J. Langley daryti išvadą, kad ląstelių membranoje yra specializuotų receptorių vietų, sąveikaujančių su vaistais.
Tačiau prioritetas kuriant vaistų veikimo receptorių teoriją priklauso vokiečių fiziologui P. Ehrlichui, kuris 1906 m. Pirmą kartą įvedė terminą „receptoriai“ ir suformulavo postulatą „vaistas neveikia, jei jis nėra pritvirtintas prie ląstelės membranos“. Remiantis P. Ehrlicho teorija, vaisto molekulė turi dvi funkciškai aktyvias grupes, iš kurių viena užtikrina jos fiksaciją ant ląstelės paviršiaus narkotiko receptoriaus srityje, o antroji funkcinė grupė sąveikauja su receptoriais ir sužadina sudėtingą biocheminių reakcijų grandinę, keičiančią jo (ląstelės) fiziologinį aktyvumą. .
Taigi, jau XX amžiaus pradžioje. paaiškėjo, kad yra bent dvi interoreceptorių klasės: sensoriniai receptoriai, perduodantys informaciją apie vidaus organų ir kūno audinių būklę į centrinę nervų sistemą; žymėti receptorius, kurie sąveikauja su vaistais, kurie keičia tikslinių ląstelių funkcinį aktyvumą.
Iškart reikia pažymėti, kad ateityje vadovėlyje tekste reikia vengti painiavos terminologijoje, vaistų receptoriuose ir biologiškai aktyviose medžiagose, t. etiketėmis, arba citoreceptoriai. bus žymimi terminu „receptoriai“, tuo tarpu sensoriniai interoreceptoriai bus žymimi terminu, apibūdinančiu jų funkcinį aktyvumą, pavyzdžiui, „baroreceptoriai“, „skausmo receptoriai“ ir kt.
P. Ehrlicho atradimas vaistų receptorių ląstelių membranoje buvo atskaitos taškas plėtojant farmakologinį mokslą, ypač farmakodinamiką, kurio vienas iš pagrindinių uždavinių yra ištirti vaistų receptorius veikiančius mechanizmus.
Šiuo metu yra atskleista daugybė ląstelių receptorių, tam tikrų biologiškai aktyvių junginių sąveikos su jais ypatumai, kurie leido, viena vertus, suprasti žinomų vaistų veikimo mechanizmą, kita vertus, tai buvo pagrindas sukurti naujus labai efektyvius vaistus.
Natūralu, kad sunku įsivaizduoti, kad evoliucijos metu žmogaus kūne susiformavo įvairių sintetinių (chemiškai gautų) vaistų receptoriai, juo labiau, kad didžioji dauguma šiuolaikinėje farmacijos rinkoje pateikiamų vaistų buvo susintetinta per pastaruosius 50 ar mažiau metų. Įrodyta, kad ląstelės receptorinis aparatas yra labai senas funkcinis-struktūrinis darinys. Taigi, a- ir β-adrenerginiai receptoriai (receptoriai, kurių sąveika su norepinefrinu ir adrenalinu daro įtaką ląstelės funkciniam aktyvumui) randami ne tik gyvūnų ląstelėse, bet ir augalų ląstelių membranose, pavyzdžiui, augalo nitrozos ląstelėse, kur adrenorecentoriai reguliuoja protoplazmos (ląstelių turinio) judėjimą.
Tuomet kokie yra P. Ehrlicho atrastų vaistų receptoriai ir kodėl jie su jais sąveikauja?
Šiuo metu nekyla abejonių, kad vadinamieji vaistų receptoriai iš tikrųjų yra endogeninių (organizme gaminamų) biologiškai aktyvių medžiagų, dalyvaujančių vidaus organų ir kūno audinių funkcinio aktyvumo reguliavime, receptoriai. Tokie biologiškai aktyvūs junginiai apima medžiagas, kurios nervinio signalo perdavimo metu išsiskiria iš nervų galūnių, taip pat hormonus, vitaminus, amino rūgštis ir kt. Kiekvienai endogeninei biologiškai aktyviajai medžiagai yra nustatyti griežtai specifiniai jos receptoriai. Taigi, pavyzdžiui, organizme pagaminta biologiškai aktyvi medžiaga adrenalinas gali suaktyvinti griežtai specifinius a- ir β-adrenoreceptorius, o gliukokortikosteroidai - antinksčių žievės hormonai - sąveikauja tik su griežtai jiems būdingais gliukokortikosteroidų receptoriais.
Sintetiniai vaistai, kurie savo poveikį realizuoja sąveikaudami su ląstelės receptorių aparatais, savo chemine struktūra yra daugiau ar mažiau panašūs į endogeninius biologiškai aktyvius junginius, kurie sąveikauja su panašiais receptoriais. Pavyzdžiui, sintetiniai vazokonstriktoriai (sukeliantys kraujagyslių susiaurėjimą) narkotikai fenilefrinas yra panašūs į endogeninę biologiškai aktyvią medžiagą norepinefriną, todėl, kaip ir norepinefrinas, turi savybę stimuliuoti a-adrenoreceptorius.
Kartais dėl savo cheminės struktūros ypatumų vaistai gali sąveikauti ne su pačiu receptoriumi, o su gretima ląstelės membranos dalimi. Kadangi šiuo atveju vaistas sąveikauja ne su pačiu receptoriumi, o su gretima ląstelės membranos dalimi, jie kalba ne apie jaudinantį ar blokuojantį poveikį receptoriui, bet apie alosterinį (iš graikų alios - dar vienas, kitoks) poveikį ar poveikį. Dėl to gali pasikeisti ir membranos, esančios greta receptoriaus, struktūra, ir atskiri paties receptoriaus komponentai, o tai gali pakeisti receptoriaus jautrumą jam specifiškai biologiškai aktyviajai medžiagai. Tais atvejais, kai padidėja receptoriaus jautrumas biologiškai aktyviajai medžiagai, kalbama apie receptoriaus sensibilizaciją (iš lotynų sensus - sens) arba sensibilizaciją (iš lotynų sensibilis - jautrumas), o tais atvejais, kai receptoriaus jautrumas sumažėja, kalbama apie desensibilizaciją. receptorius.
Allosterinio efekto ypatumas slypi tame, kad vaistai, turintys tokį veikimo mechanizmą, tiesiogiai nedaro įtakos nervinio impulso perdavimui, bet modifikuoja jį norima linkme. Pavyzdžiui, anksiolitikų (vaistų nuo nerimo; sinonimas: trankvilizatoriai), kurie pagal savo cheminę struktūrą yra benzodiazepino dariniai, veikimo mechanizmas pagrįstas postsinapsinių benzodiazepinų receptorių allosterinio sužadinimo reiškiniu. Pastarųjų sužadinimas savo ruožtu skatina slopinančių gama-aminosviesto rūgšties postsinapsinių receptorių (GABA receptorių) aktyvaciją, kuris kliniškai pasireiškia pašalinant neurozinių ligų simptomus, tokius kaip nerimas, nerimas, baimė ir kt.
Receptoriai, sąveikaudami su kuriais biologiškai aktyvi medžiaga ar vaistas bet kokiu būdu keičia tikslinės ląstelės funkcinę būklę, vadinami specifiniais.
Be specifinių receptorių, išskiriami vadinamieji vaistui būdingi receptoriai. Specializuotoje medicinos literatūroje šie receptoriai dar vadinami vaistų „praradimo vieta“. Kreipdamiesi į tokius receptorius, vaistai neturi jokio biologinio poveikio, tačiau patys tampa biologiškai neaktyvūs. Šio tipo receptorių pavyzdys gali būti receptoriai, esantys ant plazmos baltymų, ypač ant vandenyje tirpių baltymų - albumino. Šio reiškinio reikšmė bus išsamiai aptariama žemiau (žr. T. 1, p. 72).
Receptorių struktūra yra gana sudėtinga, tačiau dauguma jų yra baltyminės makromolekulės arba glikoproteinai, kurie taip pat gali apimti jonus, lipidus, nukleorūgštis ir kt. Receptorius t.y. ją sudaranti baltymo makromolekulė pasižymi specifiniu, kiekvienam receptoriui būdingu, jo cheminių grupių erdviniu išdėstymu. Baltymo makromolekulės, sudarančios receptorius, gali būti integruotos (panardintos) į citoplazminės membranos lipidų dvisluoksnį arba lokalizuotos ląstelės viduje. Pagrindinė ląstelės receptoriaus funkcija yra „atpažinti“ cheminį signalą, perduodamą į jį per endogeninę biologiškai aktyvią medžiagą ir (arba) vaistus, ir paversti jį atitinkamu ląstelės biocheminiu ir (arba) biofiziniu atsaku.
Anksčiau buvo manoma, kad vaistai ar endogeninės biologiškai aktyvios medžiagos sąveikauja su „rakto ir užrakto“ tipo receptoriais, t. receptorius turi tokią struktūrą, kuri leidžia vaistui surasti „savo“ receptorių, prisijungti prie jo ir, tarsi, „įjungti“ ir „išjungti“. Tačiau tobulėjant medicinos mokslui tapo akivaizdu, kad ego nėra toks. Šiuo metu tarpląstelinių signalų virsmo ląstelėse, reguliuojančių ląstelių funkciją, molekuliniai procesai jau yra gana gerai ištirti. mechanizmai, sukeliantys endogeninių biologiškai aktyvių medžiagų ar vaistų sąveiką su receptoriais.
Sąveikaujant su endogeninės biologiškai aktyvios medžiagos ir (arba) veikiančio L C receptoriais, įvyksta konformacija - baltymo makromolekulės formos erdvinis pasikeitimas, kuris yra įvairių tarpląstelinių procesų, kurie lemia tikslinės ląstelės atsaką į tarpininką ir (arba) vaistą, sukėlėjas. Pavyzdžiui, suaktyvinus bronchų lygiųjų raumenų adrenerginius receptorius veikiant β 2 -adrenostimuliatoriaus fenoteroliui, padidėja fermento adenilato ciklazės aktyvumas, kuris prisideda prie ciklinio adenozino monofosfato (cAMP) kaupimosi ląstelėje ir dėl to ląstelių atsipalaidavimo.
Apskritai, ląsteliniai receptoriai gali būti laikomi griežtai specializuotais ląstelių „jutimo organais“, per kuriuos jie suvokia „informaciją“, kilusią, pavyzdžiui, iš centrinės nervų sistemos ir (arba) endokrininės sistemos. Nepaisant svarbaus receptoriaus aparato vaidmens, receptoriai užima tik nereikšmingą ląstelės membranos dalį. Pvz., Ląstelės M cholinerginių receptorių aparatas užima ne daugiau kaip 1/6 000 jo paviršiaus ploto.
Narkotikų sąveikos su receptoriumi savybių tyrimas, viena vertus, leidžia suprasti jo veikimo molekulinio mechanizmo pagrindus, kita vertus, suteikia informacijos apie tai, kokie turėtų būti pokyčiai vaistų struktūroje, siekiant sustiprinti jo gebėjimą sąveikauti su šiuo receptoriumi, t. . leidžia tikslingai sintetinti naujus labai efektyvius vaistus.
Fiziologinėmis sąlygomis skirtingi ląstelių receptoriai neveikia savarankiškai, bet yra nuolat sąveikaujantys tarpusavyje, taip reguliuodami specifinį ląstelės aktyvumą. Pavyzdžiui, širdies β-adrenerginių receptorių aktyvinimas endogeniniu norepinefrinu ypač sukelia širdies susitraukimų skaičiaus padidėjimą, o širdies ląstelių M-chinerinerinių receptorių aktyvacija endogeniniu acetilcholinu, priešingai, sukelia širdies susitraukimų skaičiaus sumažėjimą.
Didelis indėlis į vaistų veikimo receptorių mechanizmo supratimą buvo padarytas atradus pre- ir postsinapsinius receptorius. Sinapsė (iš graikų sinapsio - jungtis, jungtis) yra specializuota kontaktinė zona tarp nervų ląstelių ar kitų sužadinamų kūno struktūrų, užtikrinanti gaunamos informacijos perdavimą ir jos informacinės reikšmės išsaugojimą. Sinapsių struktūros ir funkcinio vaidmens tyrimas buvo pradėtas XIX amžiaus pabaigoje. po to ispanų histologas S. Ramon n Cajal (S. Ramonas prie Cajal) pasiūlė specializuotą perdavimo sistemą centrinėje nervų sistemoje. Sinapsės gavo savo vardą 1897 m., Kai anglų fiziologas C. Sherrington pasiūlė šį terminą nurodyti kontaktinę vietą tarp nervų ląstelių.
Šiuo metu yra trijų rūšių sinapsės:
1) „elektrinės“ sinapsės, kai informacija perduodama perduodant elektrinį signalą iš priešsinapsinės membranos. Šis sinapsių tipas vadinamas efaps (iš graikų kalbos. Ephapsis - tankus kontaktas);
2) „cheminės“ sinapsės, kurių metu informacija perduodama per specialias biologiškai aktyvias medžiagas - neuromediatorius (iš graikų kalbos. Neuronas - nervas ir lotynų. Tarpininkas - tarpininkas; sinonimas: tarpininkas);
3) „mišrios“ sinapsės, kurių metu informacija perduodama tiek chemiškai, tiek elektra.
Didžiausios daugumos vaistų, turinčių įtakos sinapsių funkcijoms, farmakologinis poveikis yra realizuojamas jų poveikiu gotui ar kitam signalo perdavimo etapui cheminėse sinapsėse, t. antros rūšies sinapsėse.
Paprastai chemines sinapses neurotransmiteriai, perduodantys jose nervinius impulsus, klasifikuoja taip:
Sinapsės, kuriose acetilcholinas veikia kaip tarpininkas, vadinamos cholinerginėmis;
Sinapsės, kuriose adrenalinas ir norepinefrinas veikia kaip tarpininkas, vadinamos adrenerginėmis;
Sinapsės, kuriose ATP ir adenozinas veikia kaip tarpininkas, vadinamos purinerginėmis;
Sinapsės, kuriose gama-aminosviesto rūgštis veikia kaip tarpininkė, vadinamos GABA-erginėmis ir kt.
Šiuo metu sinapsių struktūra yra gerai suprantama. Sinapsę sudaro presinapsinis nervinės ląstelės procesas (aksonų galas) ir „signalą“ gaunantis aparatas, esantis efektoriaus („vykdomosios“) ląstelės membranoje.
Eferentinio neurono aksonas, artėdamas prie efektorinės ląstelės, praranda mielino apvalkalą, išsiplečia ir suformuoja vadinamąjį presinapsinį sustorėjimą (1.5 pav.). Nervinio paviršiaus, nukreipto į efektorinės ląstelės membraną, paviršius vadinamas presinapsine membrana. Efektoriaus ląstelės vieta, esanti prieš presinapsinę membraną, vadinama postsinapsine membrana (žr. 1.5 pav.). Priklausomai nuo sinapsės struktūrinių ypatybių, presinapsinė membrana gali turėti daugiau ar mažiau raukšlių, taigi jos plotas yra didesnis ar mažesnis. Cheminėse sinapsėse presinapsinė membrana tiesiogiai nesiliečia su postsinapsine membrana, bet yra atskirta nuo jos nedideliu atstumu, vadinamu sinapsiniu plyšiu (žr. 1.5 pav.).
Presinapsinis sustorėjimas, t. galinėje aksono dalyje yra didesnis mitochondrijų, tarpląstelinių organelių, dalyvaujančių energijos sintezėje ir kaupime, skaičius, didesnis nei neurono kūnas, o tai rodo šiame nervų ląstelės skyriuje vykstančių energetinių procesų intensyvumą. Presinapsiniuose sustorėjimuose, be mitochondrijų, yra daugybė mažų pūslelių - pūslelių. Viename presinapsiniame sustorėjime vidutiniškai yra apie 20 000 pūslelių. Pastarosios yra nevienodai išdėstytos presinapsiniame sustorėjime, kaip taisyklė, dauguma jų yra šalia presinapsinės membranos. Neuromediatorius sintetinamas neurono kūne ir aksonuose ir kaupiasi pūslelėse. Kiekvienoje pūslelėje yra keli tūkstančiai neurotransmiterio molekulių (nuo 1 000 iki 50 000). Kai atsiranda nervinis impulsas, pūslelė susilieja su presinapsine membrana, o neuromediatorius išskiriamas į sinapsinį plyšį (žr. 1.5 pav.).
Fig. 1.5. „Cheminės“ sinapsės struktūros schema:
a yra scheminis vaizdas; b - elektroninis mikrografas; 1- presinapsinis nervo galas; 2 - prssinapsinė membrana; 3 - postsinapsinė membrana; 4 - sinapsinis plyšys; B - pūslelė; NM - neurotransmiteris; P - postsinapsinis receptorius: OZ - "atvirkštinis" neurotransmiterio gaudymas; SF yra specializuotas fermentas, kuris sunaikina perteklinį neuromediatorių sinapsiniame plyšyje
Funkcionaliai aktyvios receptorių formacijos yra ant postsinapsinės membranos, sugebančios sąveikauti su neuromediatoriumi, išlaisvintu iš presinapsinės membranos, praeinant nerviniam impulsui. Receptoriai, esantys ant postsinapsinės membranos, specializuotoje medicinos literatūroje vadinami sinapsiniais arba postsinapsiniais receptoriais. Postinapsiniais receptoriais vadinamos baltyminės prigimties makromolekulės, integruotos į postsinapsinę membraną, turinčios genetiškai iš anksto apibrėžtą struktūrą ir funkcijas, galinčios grįžtamai sąveikauti su neurotransmiteriais ir (arba) vaistais dėl aktyviojo centro funkcinių grupių („atpažįstančios“ makromolekulės dalį).
Nervinio signalo perdavimas sinapsėje vyksta taip: veikiant nerviniam dirgikliui, pūslelės juda į presinapsinę membraną, o neurotransmiteris eksocitozės būdu išskiriamas į sinapsinį plyšį (žr. 1.5 pav.). Į sinapsinį plyšį išleistas neuromediatorius pasiekia postsinapsinę membraną, kur, sąveikaudamas su postsinapsiniu receptoriumi, jis sukelia biocheminių ir (arba) biofizinių reakcijų grandinę, kurios rezultatas yra fiziologinė tikslinės ląstelės reakcija. Tačiau ne visas išskiriamas neuromediatorius pasiekia postsinapsinius receptorius ir sąveikauja su jais. Dalį neuromediatorių užfiksuoja presinapsinė membrana ir „grįžta“ į laikymo vietas. Šis reiškinys vadinamas neurotransmiterių reabsorbcijos reiškiniu.
Likusį nesąveikaujančių neuromediatorių receptorių kiekį sinapsiniame plyšyje sunaikina specializuoti fermentai. Šis reiškinys vadinamas neurotransmiterių skilimu. Pvz., Fermentas acetilcholinesterazė katalizuoja (pagreitina) skaidymo (sunaikinimo) procesą neuromediatoriaus acetilcholino sinapsiniame plyšyje.
Skirtingai nuo neuromediatoriaus, jo metaboliniai produktai turi neurotransmiterio aktyvumą. Visas neurotransmiterio sąveikos su receptoriais procesas ir jo pertekliaus sunaikinimas specifiniu fermentu yra labai trumpas ir neviršija 2 ms (1 ms \u003d 0,001 s).
Tokia trumpa šio proceso trukmė, viena vertus, paaiškinama ypač greitu neurotransmiterio išsiskyrimu iš receptorių, kita vertus, dideliu fermentinio neurotransmiterio inaktyvacijos greičiu sinapsinėje šelfe.
Iš esmės sinapsės funkcinį aktyvumą galima pakeisti taip:
Pagreitinti, sumažinti arba blokuoti presinapsiniame neurotransmiterio sintezę, kaupimąsi ir (arba) katabolizmą (sunaikinimą). Dėl to kažkaip pasikeis neuromediatoriaus turinys ir jo fiziologinio aktyvumo intensyvumas.
Pavyzdžiui, simpatolitinis rezerpinas neleidžia katecholaminams kauptis sinapsinėse pūslelėse iki visiško jų ištuštinimo. Dėl to į sinapsinį plyšį išleisto neuromediatoriaus norepinefrino kiekis smarkiai sumažėja. Sistemos lygiu šis poveikis pasireiškia sumažėjus kraujospūdžiui. Kai kurie vaistai tiesiogiai nedaro įtakos presinapsinių neuromediatorių turiniui, tačiau slopina juos naikinančių fermentų veiklą. Taigi veikia nemažai antidepresantų. Pavyzdžiui, antidepresantas pirlindolis slopina (slopina) fermento monoaminooksidazės aktyvumą presinapsiniame terminacijoje ir dėl to padidina neurotransmiterių, tokių kaip norepinefrinas, dopaminas ir serotoninas, koncentraciją jame. Kliniškai šis pirlindolio poveikis pasireiškia sumažėjusiu nerimo ir baimės jausmu, pagerėjusia nuotaika, padidėjusiu fiziniu aktyvumu ir kt .;
Pakeiskite (palengvinkite, apsunkinkite) neurotransmiterio sugebėjimą prasiskverbti į presinapsinę membraną, todėl padidinkite arba sumažinkite į sinapsinį plyšį išleidžiamo neurotransmiterio kiekį kiekvienu impulsu.
Pavyzdžiui, psichostimuliatorius amfetaminas palengvina katecholaminų išsiskyrimą adrenerginėse centrinės nervų sistemos sinapsėse ir taip padidina jų kiekį sinapsiniame plyšyje. Kliniškai šis vaisto poveikis pasireiškia pagerėjusia nuotaika, padidėjusio jėgos pojūčio padidėjimu. Stabligės toksinas blokuoja slopinančių neurotransmiterių (GABA, glicino) išsiskyrimą centrinėje nervų sistemoje ir taip smarkiai sumažina jų kiekį sinapsiniame plyšyje, kuris kliniškai pasireiškia traukulių išsivystymu;
Blokuokite arba stimuliuokite neuromediatorių reabsorbciją presinapsinėje membranoje, taigi padidinkite arba sumažinkite neuromediatorių koncentraciją sinapsiniame plyšyje.
Pavyzdžiui, triciklis antidepresantas imipraminas blokuoja neuromediatoriaus norepinefrino pasisavinimą presinapsinėje membranoje ir taip smarkiai padidina jo koncentraciją sinapsiniame plyšyje. Kliniškai šis imipramino poveikis pasireiškia pagerėjusia nuotaika, padidėjusia psichine ir fizine veikla;
Stimuliuokite arba blokuokite fermentų, kurie sunaikina neuromediatorių sinapsiniame plyšyje, veiklą.
Pavyzdžiui, anticholinesterazės vaistas fizostigminas sumažina fermento acetilcholinesterazės, kuris naikina neuromediatorių acetilcholiną sinapsiniame plyšyje, aktyvumą ir taip prisideda prie jo koncentracijos padidėjimo, kuris gali pasireikšti kliniškai, visų pirma, mažinant akispūdį ir vyzdžio susiaurėjimą.
Stimuliuoja arba blokuoja postsinapsinius receptorius, t. imituoja ar blokuoja neuromediatorių poveikį.
Pavyzdžiui, narkotiniai analgetikai, kurie sužadina postsinapsinius opioidinius receptorius ir tokiu būdu imituoja neuromediatorių - enkefalinų - poveikį. Strychninas, blokuodamas slopinančio neurotransmiterio glicino receptorius, trukdo realizuoti jo slopinamąjį poveikį, todėl didelėmis dozėmis vartojamas strychninas sukelia traukulius.

Fig. 1.6. Pre-ir postsinapsinių receptorių lokalizacijos schema, parodyta adrenerginėje sinapsėje (paaiškinimas
NM - neurotransmiteris; M 2 (-) - cholinerginį „slopinantį“ presinapsinį heteroreceptorių; β 1 (+) - adrenerginiai presinapsinius „aktyvinančius“ autoreceptorius; β - adrenerginiai postsinapsiniai receptoriai
Be receptorių, esančių ant postsinapsinės membranos, t. postsinapsiniai receptoriai, receptoriai, esantys ant presinapsinės membranos, t. presinatiniai receptoriai (1.6 pav.). Nepaisant to, kad tiek pre-, tiek postsinapsinius receptorius gali sujaudinti tas pats neuromediatorius, šių receptorių formacijų funkcinis vaidmuo sinapsėse yra skirtingas. Jei postsinapsiniai receptoriai yra galutinė grandis, perduodanti nervinį impulsą į efektorinį organą, t. užtikrina vienpusį nervinio impulso laidumą iš centro į periferiją, tada presinapsiniai receptoriai dalyvauja
sinapsės neurotransmiterio aktyvumo reguliavimas, t. tam tikru mastu turi įtakos neurotransmiterio išsiskyrimo ir (arba) sintezės procesams jame. Reikia pabrėžti, kad presinapsiniai receptoriai tiesiogiai nedalyvauja atliekant nervo impulsą iš neurono į efektorinį organą.
Presinapsiniai receptoriai yra suskirstyti į dvi dideles grupes: auto- ir heteroneuromoduliacinius receptorius (žr. 1.6 pav.).
Presinapsiniai autoreceptoriai apima receptorius, kuriuos jaudina jų pačių siųstas neuromediatorius.
Pavyzdžiui, sinapsėse, esančiose sąlytyje tarp somatinių nervų ir susiaurėjusio raumens, kai sinapsiniame plyšyje yra perteklius acetilcholino, sąveikaujant su presinapsiniais autoreceptoriais, jis slopina naujos neurotransmiterio dalies išsiskyrimą. presinapsinių autoreceptorių sužadinimas reguliuoja acetilcholino išsiskyrimą iš presinapsinių gnybtų.
Tačiau ant presinapsinės membranos, be autoreceptorių, t. receptoriai, kurie yra jautrūs neuromediatoriui, kuris perduoda sužadinimą tam tikroje sinapsėje, receptoriai gali būti tokie, kurie nėra jautrūs neuromediatoriui, kuris perduoda sužadinimą tam tikroje sinapsėje, bet kurie sąveikauja su kito tipo neurotransmiteriu.
Pvz., Ant sinapsinės presinapsinės membranos, kurioje neurotransmiteris yra acetilcholinas, gali būti presinapsiniai receptoriai, jautrūs neurotransmiteriui norepinefrinui. Šio tipo presinapsiniai receptoriai vadinami heteroneuromoduliuojančiais receptoriais.
Taigi sinazė yra sudėtingas anatominis ir funkcinis darinys, užtikrinantis nervinio impulso perdavimą iš neurono į neuroną arba iš neurono į efektorinę ląstelę.
Sinapsės funkcinio aktyvumo seka (sinapsinio perdavimo stadijos) yra tokia:
Neirotransmiterio sintezė ir kaupimasis pūslelėse, lokalizuotuose presinapsiniuose sustorėjimuose (neurotransmiterio sintezė vyksta ne tik presinapsiniuose sustorėjimuose, bet ir neuronuose bei aksonuose);
Neurotransmiterio išsiskyrimas į sinapsinį plyšį, kai praeina nervinis impulsas;
Neurotransmiterio sąveika su postsinapsiniais receptoriais, kuris reiškia receptorių aktyvaciją ir efektorinės ląstelės funkcinio aktyvumo pasikeitimą;
Neurotransmiterio (fermentinio) inaktyvacija ir (arba) jo reabsorbcija presinapsinėje membranoje, t. sinapsės sugebėjimo vėl perduoti nervinį impulsą į efektorinę ląstelę atkūrimas.
Sinapsės turi šias pagrindines savybes:
Vienašalis sužadinimo laidumas (nervinis impulsas gali praeiti tik nuo presinapsinės membranos iki postsinapsinio);
Sinapsinis vėlavimas, t. tam tikras laikas praleidžiamas nervinio impulso perdavimui sinapsėje. (Sinapsių perdavimo greitis vidutiniškai yra daugiau nei 10 kartų mažesnis nei nervo impulso plitimo per nervą greitis. Cheminės sinapsės atveju jis paprastai svyruoja nuo 0,2–0,5 ms);
Nuovargis - laipsniškas nervinio impulso perdavimo sumažėjimas arba visiškas nutraukimas su ilgai trunkančia nervų stimuliacija. Šio reiškinio pagrindas yra, viena vertus, neuromediatorių atsargų išeikvojimas presinapsiniuose sustorėjimuose, ir, kita vertus, jautrumo sumažėjimas neurotransmiterio postsinapsiniams receptoriams;
Didelis sinapsinių formacijų jautrumas vaistams ir nuodams.
Būtent paskutinė sinapsių savybė yra pagrįsta vaistų, veikiančių sinapsių, esančių įvairiuose kūno organuose ir audiniuose, funkcinei veiklai, farmakologija. Reikia pabrėžti, kad farmakologinio poveikio objektas gali būti bet kuri sinapsinio perdavimo stadija. Kaip vaistai, turintys įtakos sinapsiniam perdavimui, naudojami egzogeniniai neuromediatorių analogai, jų cheminiai pirmtakai ir kitos biologiškai aktyvios medžiagos, kurios bet kokiu būdu gali pakeisti sinapsės funkcinį aktyvumą.
Reikėtų pažymėti, kad daugelis vaistų turi ne vieną, o kelis efekto taikymo taškus sinapsių lygmenyje. Taigi, pavyzdžiui, antidepresantas pirlindolis ne tik slopina monoaminooksidazės fermento aktyvumą sinapsiniame plyšyje, bet ir blokuoja norepinefrino pasisavinimą presinapsinėje membranoje.
Kalbant apie receptorių lokalizaciją sinapsėse, juos galima suskirstyti į presinapsinius, postsinapsinius ir ekstrasinapsinius. Pastarieji, pavyzdžiui, apima receptorius, esančius ant trombocitų ląstelių membranų.
Ląstelių topografijos (vietos) požiūriu receptorius taip pat galima klasifikuoti pagal jų vietą ląstelinėse struktūrose:
membraniniai receptoriai - receptoriai, esantys ant citoplazminės membranos;
citozoliniai receptoriai - receptoriai, esantys ant ląstelių formacijos;
branduoliniai receptoriai - receptoriai, esantys ant ląstelės branduolio membranos.
Kaip minėta anksčiau, dėl sąveikos su endogeninių biologinių medžiagų ar vaistų receptoriais keičiasi tikslinių ląstelių funkcinis aktyvumas. Šis procesas gali būti įgyvendinamas skirtingais būdais, griežtai apibrėžtais skirtingų tipų receptoriais. Remiantis tuo, šiuo metu yra išskiriami keturi receptorių tipai, kurių kiekvienas turi savo, iš esmės skirtingą nuo kitų, mechanizmą, kuriuo signalas iš receptoriaus inicijuoja biocheminių ir (arba) biofizinių reakcijų kaskadą, dėl kurios keičiasi tikslinių ląstelių funkcinė būklė.
Pirmieji trys receptorių tipai yra lokalizuoti ląstelių (citoplazminėje) membranoje, o ketvirtasis receptorių tipas apima citozolinius ir branduolinius receptorius.
I tipo receptoriai apima ląstelinius (membraninius) receptorius, kurie savo poveikį realizuoja per vadinamuosius signalinius G baltymus (1.7 pav.).
Pirmajame etape biologiškai aktyvi medžiaga ar vaistas, „einantis“ link ląstelės membranos, „atpažįsta“ receptorių ir sąveikauja su juo, o po to receptorius suaktyvina specializuotą signalo G baltymą, esantį vidiniame membranos paviršiuje. Be to, aktyvuotas G baltymas keičia vidinio efektoriaus elemento, kuris, kaip taisyklė, yra fermentai, funkcinį aktyvumą. Tada efektorinis elementas, kuris yra fermentas, suaktyvina antrinį pasiuntinį arba antrinį pasiuntinį, kuris suaktyvina biocheminių reakcijų, pakeičiančių tikslinių ląstelių funkcinį aktyvumą, kaskadą.
I tipo ląstelių receptoriai, t. receptoriai, konjuguoti su signalo G baltymais, yra struktūriškai panašūs vienas į kitą, o pagal savo erdvinę struktūrą yra serpentino (iš prancūzų serpantino - gyvatės, rutulio) struktūros (1.8 pav.).
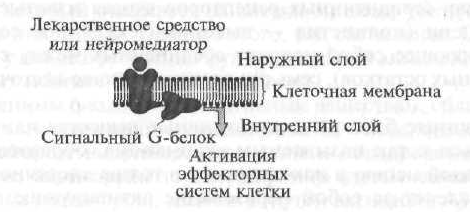
Fig. 1.7. 1 tipo receptorių struktūra (paaiškinimas tekste)
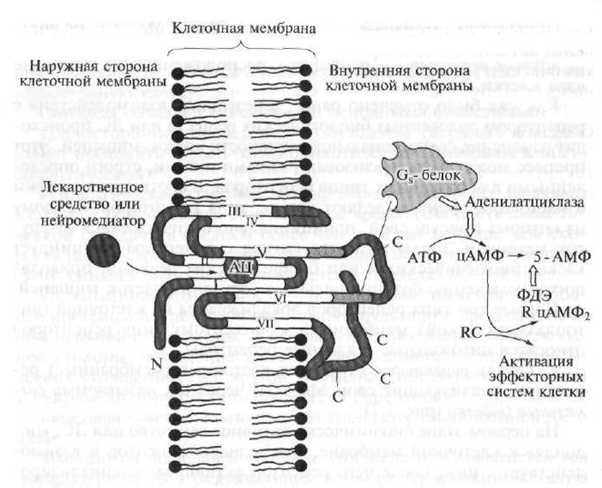
Fig. 1.8. "Serpentino" struktūros schema
receptorius:
N yra polipeptido receptoriaus dalis, esanti virš ląstelės membranos; C yra polipeptido receptoriaus dalis, esanti po ląstelės membrana; AC - aktyvusis receptoriaus, su kuriuo vaistas sąveikauja, centras; eina neurotransmiteris; ATP - adenozino trifosfatas - antrinis pasiuntinys; cAMP - ciklinis adenozino monofosfatas; 5-AMP - adenozino-5 "monofosfatas; PDE - fosfodiesterazė; R. RC - nuo cAMP priklausomas fermentas (baltymų kinazė) su reguliuojančiais ir kataliziniais (pagreitinančiais reakciją) subvienetais; 1-VII - serpentino receptorių polipeptidinėmis grandinėmis
Serpentino receptorius sudaro sukinamos polipeptido grandinės (polipeptidas yra didelės molekulinės masės junginys, kuris yra sujungtų aminorūgščių liekanų grandinė), kurie septynis kartus prasiskverbia į ląstelės membraną.
Endogeninės biologiškai aktyvios medžiagos ar vaistai gali jungtis prie vadinamosios „kišenės“, kurią suformuoja polipeptido grandinė ir esanti ląstelės membranos storyje, o tai reiškia, kad susidaro aktyvinantis signalas, kuris perduodamas į receptorių grandinės dalis, esančias ląstelės citoplazmoje. Signaliniai G baltymai sąveikauja su
stolinės (tarpląstelinės) grindų ir peptidų grandinės dalys; suaktyvinti ir paleisti biocheminių reakcijų kaskadą, kuri keičia jos funkcinį aktyvumą tikslinės ląstelės atžvilgiu, t. inicijuoti pirminį farmakologinį atsaką.
Šiuo metu žinomi keli signalinių G baltymų tipai.
G signalas, -baltymai. Šie signaliniai baltymai, kaip taisyklė, aktyvina efektorinį elementą - fermento adenilato ciklazę, kuri savo ruožtu stimuliuoja antrinio pasiuntinio - ciklinio adenozino monofosfato (cAMP) - sintezę ląstelėje (iš ATP). Biologinis cAMP, kaip antrinio pasiuntinio, vaidmuo yra labai svarbus. Pavyzdžiui, padidėjus jo kiekiui širdies ląstelėse, padidėja širdies susitraukimų dažnis ir stiprumas. Be to, padidėjusi cAMP koncentracija įvairiose tikslinėse ląstelėse lemia kraujagyslių ir bronchų lygiųjų raumenų atsipalaidavimą, energijos atsargų mobilizavimą (angliavandenių suirimą kepenyse), slopina trombocitų agregacijos gebėjimą, mažina miometrijos (gimdos raumenų) ir šlapimo pūslės tonusą ir kt.
Daugybė neurotransmiterių, tokių kaip adrenalinas (aktyvuodami β-adrenerginius receptorius), dopaminas (aktyvuodami D 1 -dopamino receptorius), adenozinas (aktyvindami adenozino A 2 receptorius) priklauso endogeninėms biologiškai aktyvioms medžiagoms, turinčioms galimybę suaktyvinti G signalizuojančius baltymus. histaminas (aktyvinant histamino G 2 receptorius), serotoninas (aktyvinant serotonino 5-HT 4 receptorius), taip pat daugybė hormonų, pavyzdžiui, vazopresinas (stimuliuojant V 2 -vasopresino receptorius) ir kt.
Signalo G i-baltymai. Skirtingai nuo signalo G s baltymų, signalo G i baltymų aktyvacija ne stimuliuoja, bet slopina efektoriaus elemento - adenilato ciklazės fermento - aktyvumą, dėl kurio sumažėja cAMP koncentracija antrinio pasiuntinio tikslinėse ląstelėse. CAMP kiekio sumažėjimas tikslinėse ląstelėse sukelia širdies susitraukimų sumažėjimą, kraujagyslių ir bronchų tonuso padidėjimą, t. cAMP turinio padidėjimas tikslinėse ląstelėse priešingai. Be to, nemažai signalizuojančių G i baltymų dalyvauja reguliuojant transmembraninių joninių Ca 2+ ir K + kanalų funkcinį aktyvumą.
Nemažai neurotransmiterių, pavyzdžiui, adrenalinas ir norepinefrinas (suaktyvindami 2-adrenoreceptorius), dopaminas (suaktyvindami D2 - dopamino receptorius), adenozinas (suaktyvindami A 1) priklauso endogeninėms biologiškai aktyvioms medžiagoms, gebančioms suaktyvinti G i baltymus. adenozino receptoriai), acetilcholinas (aktyvinant M 2 ir M 4 muskarininius receptorius) ir kt.
Signaliniai G ^ baltymai. Šie signaliniai baltymai prisideda prie kito tikslinių ląstelių efektorinio elemento, fosforilazės C fermento, aktyvavimo, kuris savo ruožtu skatina antrinių pasiuntinių, diacilglicerolio (DAG) ir inozitol-1,4,5-trifosfato (ITP) susidarymą tikslinėse ląstelėse. Pirmasis iš jų (DAG) yra susijęs su ląstelės membrana ir inicijuoja biochemines reakcijas, susijusias su kontraktilinės būklės reguliavimu, ląstelių augimu ir dalijimusi bei tam tikrų hormonų sekrecija tikslinėmis ląstelėmis. Veikiant fermentui fosfolipazės A 2, DAG gali būti metabolizuojamas į arachidono rūgštį, kuri dalyvauja biologiškai aktyvių medžiagų, tokių kaip eikosanoidai, sintezėje - prostaglandini, prostaciklinai, tromboksanai, leukotrienai (žr. T. I, p. 478).
Antrasis antrinis pasiuntinys, ITF, nėra fiksuotas ant ląstelės membranos ir pereina į viduląstelinę terpę (citozolį), kur jis inicijuoja Ca 2+ jonų išsiskyrimą iš ląstelių saugyklų, t. skatina neaktyvių Ca 2+ jonų perėjimą į aktyviąją formą.
Daugelis tyrinėtojų mano, kad Ca 2+ jonai yra tretinis tarpininkas arba tarpininkas. Taip yra dėl to, kad Ca 2+ jonų vaidmuo reguliuojant ląstelių funkcinį aktyvumą yra labai svarbus. Ca 2+ jonai gali patekti į ląstelę iš išorinės aplinkos per specialius transmembraninius jonų kanalus ir (arba) išlaisvinti iš ląstelių saugyklų. Pagrindinis depas (neaktyvių Ca 2+ jonų kaupimosi vieta) ląstelėje yra endoplazminis, arba sarkoplazminis, retikulumas (retikulinis sarkoplazminis sinonimas, sinonimas: endoplazminis retikulumas - tarpląstelinė organelė, tai citoplazmoje esančių kanalėlių ir cisternų sistema, apribota membrana; ji dalyvauja). medžiagų gabenimas citoplazmoje). Iš sarkoplazminio retikulio į citoplazmą patenkantys laisvieji (aktyvūs) Ca 2+ jonai sąveikauja su kai kuriais Ca 2+ rišančiais baltymais, iš kurių svarbiausias yra kalmodulinas. Kompleksas „kalmodulinas-Ca 2+“ ir (arba) Ca 2+ jonų kompleksai su kitais kalcį jungiančiais baltymais sukelia ląstelėje biocheminių reakcijų kaskadą. Dėl to, priklausomai nuo taikinių organų, kuriuose vyksta šis procesas, padidėja miokardo ir griaučių raumenų susitraukimo funkcija, padidėja kraujagyslių, bronchų ir gimdos lygiųjų raumenų tonusas, padidėja liaukinio audinio sekrecinis aktyvumas, stimuliuojamas neuromediatorių išsiskyrimas iš nervų galūnių ir kt. . Taip pat įrodyta, kad Ca 2+ jonai gali padidinti fermentų, dalyvaujančių baltymų, angliavandenių ir riebalų metabolizme, aktyvumą.
Be tiesioginio ryšio tarp antrinių pasiuntinių - DAG ir ITF, taigi ir signalo G q baltymų, Ca 2+ jonai fiziologinėmis sąlygomis turi gana sudėtingą sąveiką su cAMP antriniu pasiuntiniu, kurio aktyvumą reguliuoja signalo G s ir G i baltymai. Taigi buvo parodyta, kad laisvieji Ca 2+ jonai, patenkantys į nervinės ląstelės citoplazmą per kalmodulino-Ca 2+ sistemą, pradeda mažinti iAMP kiekį ląstelėje. Tuo pačiu metu, norint išlaikyti atvirą kalcio jonų kanalų būseną ląstelėje, būtinos didelės cAMP koncentracijos, t. cAMP kiekio sumažėjimas, kurį sukelia kalmodulino-Ca 2+ kompleksas, reiškia, kad reikia nutraukti laisvų Ca 2+ jonų vartojimą citoplazmoje. Kita vertus, yra įrodymų, kad cAMP antrinis pasiuntinys padidina laisvųjų Ca 2+ jonų absorbciją sarkoplazminiu retikuliu, t. skatina Ca 2+ jonų perėjimą iš laisvos, aktyvios formos į surištą, neaktyvią formą.
Padidėjus antrinių pasiuntinių kiekiui tikslinėse ląstelėse - DAG ir ITF - padidėja lygiųjų raumenų tonusas, padidėja liaukų sekrecija, palengvėja neurotransmiterių išsiskyrimas iš presinapsinių galūnių, padidėja trombocitų agregavimo galimybė ir kt.
Endogeninės biologiškai aktyvios medžiagos, turinčios galimybę suaktyvinti C q signalo baltymus, yra tokie neurotransmiteriai kaip norepinefrinas (suaktyvinant 1 adrenoreceptorius), acetilcholinas (suaktyvinant muskarininius M 1 ir M 3 receptorius), serotoninas (dėl serotonino aktyvacijos). 5-HT 2a receptoriai), histaminas (dėl histamino H 1 receptorių aktyvacijos), taip pat kitos endogeninės biologiškai aktyvios medžiagos, pavyzdžiui, bradikininas ir angiotenzinas.
Šiuo metu, be išvardytų signalinių G baltymų (G s, G |, G q), yra nustatyti ir kiti signaliniai G baltymai - G s, G i, G q, kurių fiziologinis vaidmuo vis dar nėra aiškus. Tačiau tuo pat metu yra įrodymų, kad, pavyzdžiui, signalo C o baltymas dalyvauja reguliuojant transmembraninių jonų kanalų funkcinį aktyvumą.
11 tipo receptorių funkcinis vienetas yra transmembraninis (prasiskverbiantis per visą ląstelės membranos storį) baltymas (fermentas). Pats receptorius susideda iš dviejų vienodų fragmentų, kurie vadinami monomerais. Monomerai yra išdėstyti nereikšmingu atstumu vienas nuo kito, o patį monomerą sudaro du funkciškai aktyvūs subvienetai - domenai, sujungti polipeptido segmentu, kertančiu lipidų dvisluoksnį membraną (1.9 pav.). Monomero a-subvienetas išsikiša virš išorinio membranos paviršiaus ir yra atsakingas už receptoriaus prisijungimą prie biologiškai aktyvių medžiagų, o P-subvienetas yra panardinamas į ląstelės citoplazmą.

Fig. 1.9. II tipo receptorių struktūra (paaiškinimas tekste): 1 - monomero a-subvienetas; 2 - monomero β-subvienetas
Prijungęs biologiškai aktyvią medžiagą prie receptoriaus α-subvienetų, receptoriai pasikeičia iš neaktyvios monomerinės būsenos į aktyvią dimerinę būseną, kurioje du monomerai susijungia membranos plokštumoje (žr. 1.9 pav.). Tokiu atveju stimuliuojamas citoplazminio β-subvieneto fermentinis aktyvumas, todėl tikslinėje ląstelėje pradedama kaskadinė biocheminių reakcijų, keičiančių jo funkcinę būseną, dalis.
Kaip transmembraninis fermentas, formuojantis receptorius, paprastai naudojami tokie fermentai kaip tirozino kinazė arba guanilato ciklazė.
Tirozinkinazės receptoriaus pavyzdys yra insulino receptoriai (žr. T. 1, p. 435).
Guanilato ciklazės signalo perdavimo kelias prasideda nuo receptoriaus α-subvieneto sąveikos su endogenine biologiškai aktyvia medžiaga, pavyzdžiui, su prieširdžių natriuretinį faktorių (ANF), kuris yra biologiškai aktyvi medžiaga, išskiriama prieširdžių ląstelių ir dalyvaujanti širdies susitraukimų reguliavime. Dėl šios sąveikos pasikeičia receptoriaus konfigūracija, kurią sudaro jo monomerų sujungimas į dimerą. Šis procesas suaktyvina receptoriaus fermentinę dalį, esančią jo citozoliniame β-subvienete, t. fermento guanilato ciklazės, kuris savo ruožtu skatina ciklinio guanidino-3,5 "monofosfato (cGMP) antrinio pasiuntinio koncentracijos padidėjimą tikslinėje ląstelėje. Padidėjus cGMP koncentracijai tikslinėse ląstelėse, vyksta kaskados biocheminių reakcijų, kurios keičia jų funkcinę būklę, pavyzdžiui, lygiųjų raumenų ląstelių atpalaidavimas. indai.
III tipo receptoriams priskiriami receptoriai, kurie, veikiami endogeninių biologiškai aktyvių medžiagų - neurotransmiterių, užtikrina atitinkamų jonų praėjimą per ląstelės membraną, o tai lemia jo (membranos) elektrinio krūvio (potencialo) pasikeitimą.
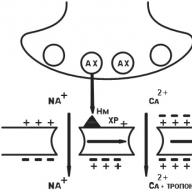
Pagal savo struktūrą III tina receptoriai žymi kanalą, prasiskverbiantį į ląstelės membranos lipidinį dvisluoksnį sluoksnį, suformuotą iš kelių polipozių (1.10 pav.). Pavyzdžiui, nikotino (H) receptorius yra kanalas, kurio skersmuo 8 nm, suformuotas iš penkių polipeptido subvienetų (a - du, β, γ, d) (žr. 1.10 pav.). Kai neurotransmiteris acetilcholinas sąveikauja su dalimi (domenu), išsikišusiu virš ląstelės membranos paviršiaus, pasikeičia receptoriaus α-subvienetas, jo struktūra ir atsidaro centrinis kanalas, per kurį Na + jonai patenka į tikslinę ląstelę pagal koncentracijos gradientą, dėl kurio keičiasi jo funkcinis veikla. Be H-chinerinerginių receptorių, gama-aminosviesto rūgšties ir sužadinamųjų aminorūgščių receptoriai yra III tipo receptoriai.
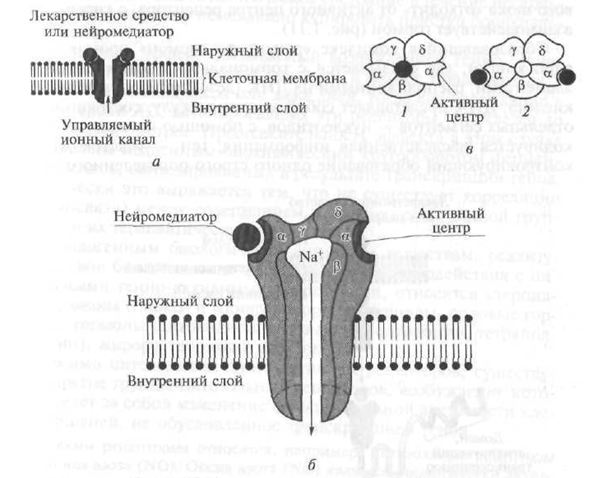
Fig. 1.10. 111 tipo receptorių struktūros schema:
a yra grandinės schema; b - transmembraninis jonų kanalas (kontekste); c - transmembraninis jonų kanalas (vaizdas iš viršaus); / - kanalas neaktyvus (uždaras); 2 - kanalas aktyviojoje (atviroje) būsenoje; a. β, γ, d - kanalo polipeptido subvienetai
IV tipo receptoriai apima tarpląstelinius ir branduolinius receptorius. Biologiškai aktyvios medžiagos, sąveikaujančios su šio tipo receptoriais, yra lipofiliniai (lengvai tirpsta riebaluose) junginiai, todėl lengvai įsiskverbia į ląstelės membraną ir pasiekia savo ląstelių receptorius. Į tarpląstelinius receptorius įeina hormonų receptoriai, taip pat kitos biologiškai aktyvios medžiagos.
Hormonų sąveikos su tarpląsteliniais receptoriais mechanizmas yra gana sudėtingas, tačiau schematiškai jį galima pavaizduoti taip. Pagal struktūrą tarpląstelinis hormonų receptorius yra polipeptidas, susidedantis iš kelių funkcinių vienetų - domenų. Trūkstant hormono, receptorius yra neaktyvus dėl to, kad jo aktyvųjį centrą blokuoja specializuotas baltymas - vadinamasis šilumos šoko baltymas. Tuo atveju, kai hormonas „artėja“ prie receptorių, šilumos šoko baltymas „nukrypsta“ nuo aktyvaus receptoriaus centro, su kuriuo hormonas sąveikauja (1.11 pav.).
Gautas receptoriaus-hormono kompleksas prasiskverbia į ląstelės branduolį, kur jis jungiasi su hormonams jautriais elementais, esančiais DNR (dezoksiribonukleorūgštis; DNR yra makromolekulė, susidedanti iš atskirų segmentų - nukleotidų, kurių pagalba genuose užkoduota paveldima informacija; genas - DNR gabalas, kontroliuojantis vieno griežtai apibrėžto baltymo susidarymą
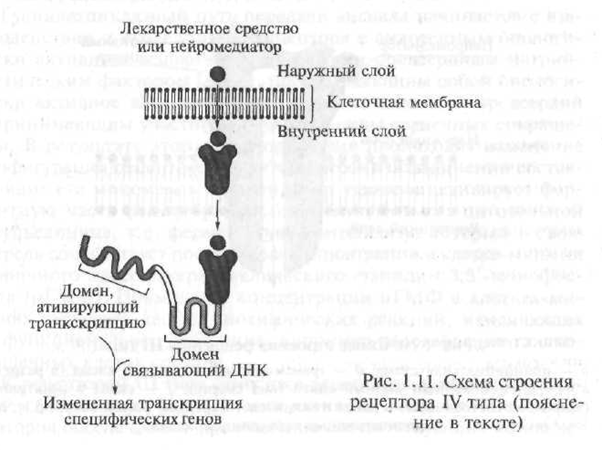
Fig. 1.11. IV tipo receptorių struktūros schema (paaiškinimas tekste)
ka). Dėl šios sąveikos pradedamas genų transkripcijos procesas - genetiniame kode esančios informacijos perkėlimas iš DNR molekulės į informacinės RNR molekulę (mRNR, sin: matricos RNR - mRNR). Transkripcija yra pirmasis baltymų susidarymo ląstelėje žingsnis. Gauta mRNR. palieka ląstelės branduolį ir juda į ribosomas - tarpląstelines organelles, atsakingas už baltymų sintezę ląstelėje. Specialiojoje medicinos literatūroje receptoriai, kurių aktyvacija sukelia genų transkripcijos procesą, yra vadinami genetiškai aktyviais receptoriais.
Paprastai tikslinių ląstelių atsakas į genetiškai aktyvių receptorių sužadinimą vystosi gana lėtai, o tai labai svarbu klinikiniu požiūriu.
Pirma, tikslinių ląstelių reakcija vėluoja laiku, nes tam reikia naujų baltymų sintezės, kuri paprastai užtrunka 20-30 minučių, t. hormonai, aktyvuodami IV tipo receptorius, per kelias minutes nesugeba pakeisti patologinės būklės, pavyzdžiui, nedelsdami sustabdo bronchinės astmos priepuolį.
Antra, genetiškai aktyvių receptorių sužadinimo poveikis yra gana ilgas ir gali trukti kelias valandas ar net dienas, o vaistų, kurie suaktyvino šiuos receptorius, kiekis kraujo plazmoje sumažėja iki nulio daug greičiau. Poveikį šiuo atveju lemia lėta fermentų ir baltymų, sintezuotų dėl genų transkripcijos, biocheminė cirkuliacija. Kliniškai tai išreiškiama tuo, kad nėra koreliacijos (ryšio) tarp tam tikros grupės vaistų plazmoje ir jų terapinio poveikio.
Endogeninėms biologiškai aktyvioms medžiagoms, kurios realizuoja savo biologinį poveikį sąveikaudamos su citozolinių genų aktyviais receptoriais, yra steroidiniai hormonai (gliuko- ir mineralokortikosteroidai, lytiniai hormonai), skydliaukės hormonai (trijodtironinas, tetrajodtironinas) ir riebaluose tirpus vitaminas D.
Be citozolinių genų aktyvių receptorių, yra ir kitų citozolinių receptorių grupių, kurių sužadinimas sukelia tikslinių ląstelių funkcinio aktyvumo pasikeitimą ne dėl genų transkripcijos.
Tokie receptoriai apima, pavyzdžiui, citozolinius azoto oksido (N0) receptorius. Azoto oksidas (N0) yra biologiškai aktyvi medžiaga, susidaranti kraujagyslių endotelyje. Kaip endogeninė biologiškai aktyvi medžiaga, azoto oksidas iš triušių uostų pirmą kartą buvo išskirtas 1987 m. Amerikiečių fiziologo R. F. Furchgotto vardu ir buvo vadinamas „endotelio atpalaiduojančiu faktoriu - ORF“. Azoto oksidas yra lipofilinis junginys, lengvai prasiskverbiantis į ląstelės membraną, kur jis sąveikauja su specifiniais citozoliniais receptoriais, o tai reiškia, kad aktyvinamas fermento guanilato ciklazė. Pastaroji, savo ruožtu, stimuliuoja cGMP antrinio pasiuntinio sintezę, sukeliančią tarpląstelinių biocheminių reakcijų kaskadą, lemiančią taikinių ląstelių, kraujagyslių lygiųjų raumenų ląstelių atsipalaidavimą.
Taigi šiuo metu yra išskiriami keturi pagrindiniai mechanizmai ir atitinkamai IV receptorių tipai dėl sąveikos su kuriomis endogeninėmis biologiškai aktyviomis medžiagomis ir (arba) jų sintetiniais analogais, t. Vaistai gali paveikti tikslinių ląstelių funkcinę būklę.
Tačiau tai nereiškia, kad žinomų biologiškai aktyvių medžiagų receptorių skaičius yra tik 4. Jie yra neišmatuojamai didesni. Taip yra dėl to, kad per tą patį pagrindinį veikimo mechanizmą labai didelis skaičius endogeninių biologinių medžiagų, turinčių įvairių cheminių struktūrų, gali įtakoti ląstelių funkcinį aktyvumą. Pavyzdžiui, neuromediatoriai norepinefrinas ir histaminas, kurie skiriasi savo chemine struktūra, todėl receptoriuose, su kuriais jie sąveikauja, perduoda sužadinimo signalą tikslinėms ląstelėms tuo pačiu pagrindiniu mechanizmu - signalo G baltymų aktyvumo stimuliavimu, t. abu sąveikauja su I tipo receptoriais.
Todėl visi šiuo metu žinomi receptoriai yra klasifikuojami remiantis ne tik signalo perdavimo į tikslinių ląstelių tarpląstelines struktūras ypatybėmis, bet ir pagal tų endogeninių biologiškai aktyvių medžiagų pavadinimus, su kuriais jie konkrečiai sąveikauja.
Reikėtų pažymėti, kad receptoriai gavo savo vardą, atsižvelgdami į endogeninių biologiškai aktyvių medžiagų, su kuriomis jie sąveikauja, pavadinimus, dar ilgai, kol tapo žinomi signalo perdavimo į tikslines ląsteles mechanizmai.
Endogeninėms biologiškai aktyvioms medžiagoms, kurios savo poveikį supranta per sąveiką su specifiniais receptoriais, yra neurotransmiteriai (acetilcholinas, norepinefrinas, dopaminas, histaminas, serotoninas ir kt.), Hormonai, audinių kilmės biologiškai aktyvios medžiagos - autocidadai (prostaglandinai, tromboksanai, leukotrienai, bradikininas, angiotenzinas ir kt.). Specialiojoje medicinos literatūroje visos šios medžiagos dažnai derinamos su terminu „ligadai“ (iš Lot. Ligo - surišti, tai yra, medžiaga, galinti surišti su receptoriais).
Taigi receptoriai gauna savo vardą iš savo specifinių ligandų pavadinimo. Pavyzdžiui, dopamino neurotransmiterio receptoriai yra vadinami dopaminu, insulino hormonu - insulinu, leukotrienų augokaitu - leukotriene ir kt.
Ateityje vadovėlio tekste, siekiant išvengti painiavos kalbant apie signalo perdavimo mechanizmą iš receptorių į viduląstelines formacijas, bus naudojamas terminas „receptoriaus tipas“, o kalbant apie receptorių pavadinimą dėl ligando sąveikos su juo bus naudojamas terminas „receptorių rūšys“.
Paprastai daugelis tos pačios rūšies receptorių yra suskirstyti į kelis potipius, pavyzdžiui, adrenerginiai receptoriai yra suskirstyti į a- ir β-adrenerginius receptorius, cholinerginiai receptoriai - į M- ir N-cholinerginius receptorius ir kt. Daugeliu atvejų porūšiai taip pat yra suskirstomi į mažesnes grupes: β 1 - ir β 2 -adrenoreceptoriai, N n - ir N m-cholinerginiai receptoriai ir kt.
Receptorių porūšių nustatymas ir mechanizmų, pagal kuriuos endogeninės biologiškai aktyvios medžiagos sąveikauja su jais, tyrimas yra labai svarbus šiuolaikinei farmakologijai, nes tai leidžia sukurti vaistus, kurie sąveikauja su griežtai apibrėžtais receptorių porūšiais. Pavyzdžiui, padalijus β-adrenerginius receptorius į β 1 (daugiausia lokalizuotas ant širdies ląstelių ląstelių membranos) ir β 2 (lokalizuotas, pavyzdžiui, ant bronchų lygiųjų raumenų ląstelių membranų), atsirado galimybė sukurti vaistus, selektyviai veikiančius širdies raumenį (β 1 -adrenostimuliatoriai). ) - vaistas nonachlazinas, selektyviai veikiantis lygiuosius bronchų raumenis (β 2 -adrenostimuliatoriai) - vaistas salbutamolis ir kt.
Reikėtų pažymėti, kad kūno organuose ir audiniuose nėra pastovaus receptorių skaičiaus ir (arba) jų porūšių, t. jis kintamas. Tiek patologiniai procesai, tiek vaistai gali pakeisti receptorių skaičių organe.
Pavyzdžiui, koronarinę širdies ligą lydi padidėjęs 3-adrenerginių receptorių skaičius širdies raumenyje, o pacientams, sergantiems hipertenzija, padidėja ir a-, ir β-adrenerginių receptorių skaičius. Antidepresantas imipraminas, ilgą laiką vartojant, sumažina β-adrenerginių receptorių kiekį smegenų audinyje. Tokių pavyzdžių yra nemažai.
Endogeninių (organizme gaminamų) neuromediatorių ar vaistų afinitetas receptoriams yra apibūdinamas terminu „afinitetas“, o jų prisijungimo prie receptorių greitis ir stiprumas žymimas terminu „afinitetas“.
Natūralu, kad vaistų sąveika su receptoriumi nėra savaiminis tikslas, tačiau tai turėtų šiek tiek pakeisti organų ar organizmo audinių veiklą.
Toks pokytis arba reakcija, atitinkanti šio receptoriaus funkcinę reikšmę, vadinama vidiniu vaistų aktyvumu.
Vaistai, turintys vidinį aktyvumą ir afinitetą receptoriams, yra agonistai, t. veikia kaip endogeninės biologiškai aktyvios medžiagos.
Pvz., Fenilefrinas, a-adrenerginių receptorių stimuliatorius, daro poveikį arteriolėms, panašioms į neurotransmiterį norepinefriną, t. jis yra adrenoreceptorių agonistas. Specialiojoje medicinos literatūroje, be termino „agonistas“, kartais vartojamas terminas „receptorių stimuliatorius“ arba „mimetikas“, pavyzdžiui, adrenomimetikas, t. Vaistai, kurie stimuliuoja adrenerginius receptorius.
Vaistai, kurie turi afinitetą receptoriams, tačiau slopina egzogeninių ir endogeninių agonistų sąveiką su receptoriais, vadinami antagonistais.
Pvz., Atropino M-cholinerginių receptorių blokatoriai trukdo acetilcholino neurotransmiterio M-cholinerginių receptorių sąveikai; blokatoriai (propranololio J adrenerginiai receptoriai, blokuojantys β 2 -adrenostruktūras plaučiuose), slopina stimuliuojantį salbutamolio β 2 -adrenostimuliatoriaus poveikį jiems, t. y. atropinas ir propranololis yra atitinkamų receptorių antagonistai.
Specializuotoje medicinos literatūroje, be termino „antagonistas“, vartojamas terminas „receptorių blokatoriai“ arba „litikai“, pavyzdžiui, anticholinerginiai vaistai, t. vaistai, blokuojantys cholinerginius receptorius.
Schema, receptorių sąveika su agonistais ir antagonistais parodyta fig. 1.12.
Agonistai gali veikti tiek tiesiogiai, tiek netiesiogiai, t. netiesioginis veiksmas.

Fig. 1.12. Receptoriaus sąveikos su agonistu (a) ir antagonistu (6) su receptoriumi schema (paaiškinimas tekste)
Pavyzdžiui, opioidų receptorių agonistas - vaistas morfinas - suvokia savo poveikį tiesiogiai stimuliuodamas (mu) p-, (kappa) k- ir (delta) 8-opioidinius receptorius, t. daro jiems tiesioginį stimuliuojantį poveikį, o simpatomimetinis efedrinas savo ląstelinį poveikį realizuoja netiesiogiai ar netiesiogiai stimuliuodamas a- ir β-adrenerginius receptorius: jis padeda išstumti neurotransmiterį norepinefriną iš presinapsinių nervų galūnių, slopina jo pasisavinimą nervų galūnėmis ir padidina a- ir p- jautrumą. adrenoreceptorių prie norepinefrino ir epinefrino ir stimuliuoja adrenalino išsiskyrimą iš antinksčių žievės. Taigi efedrinas realizuoja savo farmakologinį poveikį ne tiesiogiai, o per neurotransmiterį, tuo tarpu tiesiogiai nebendrauja su receptoriais.
Antagonistai, taip pat receptorių agonistai, gali realizuoti savo farmakologinį poveikį tiesiogiai arba netiesiogiai, blokuodami atitinkamus receptorius.
Pavyzdžiui, skrandžio H 3 histamino receptorių blokatorius ranitidinas tiesiogiai sąveikauja su skrandžio H 2 histamino receptoriais, slopina bazinę druskos rūgšties sekreciją, slopindamas sąveiką su endogeniniu histamino neurotransmiterio receptoriumi, t. tiesiogiai blokuoja receptorius, o simpatolitinis rezerpinas, patenkantis į kūną, kaip simpatomimistinis efedrinas „išstumia“ neurotransmiterį norepinefriną iš presinapsinių galūnių, tuo pačiu blokuodamas naujų norepinefrino „porcijų“ sintezę, o tai lemia greitą mediatorių atsargų išeikvojimą presynapsinėse galūnėse. Dėl to susidaro būklė, kai indų a-adrenerginiai receptoriai „neveikia“ dėl savo fiziologinio stimuliatoriaus noradrenapino trūkumo. Mes galime pasakyti, kad simpatolitinis rezerpinas sukelia funkcinę, netiesioginę adrenoreceptorių blokadą.
Kai kurie vaistai sujungia agonistų ir antagonistų savybes, t. tam tikromis sąlygomis tie patys receptoriai sužadina arba blokuoja. Tais atvejais, kai stimuliuojantis komponentas vyrauja dėl vaisto farmakologinio poveikio, jie sako, kad vaistas yra dalinis arba dalinis agonistas.
Dalinių ar dalinių agonistų veikimo mechanizmas yra tas, kad šie vaistai, artėjant prie receptoriaus, yra fiksuojami ant jo. Tačiau dėl savo cheminės struktūros ypatumų jie ją sužadina taip, kad sukelia ne visišką, o tik dalinį stimuliuojantį atsaką. Tuo pačiu metu jie trukdo kitų vaistų ir (arba) atitinkamo neurotransmiterio sąveikai su šiuo receptoriumi.
Pavyzdžiui, nalorfinas, kuris vartojamas perdozavus narkotinių analgetikų, yra dalinis agonistas. Nalorfinas, „einantis“ į opioidų receptorius, išstumia morfiną iš jo jungties, ty sustabdo agonistinį morfino poveikį receptoriui. Nedidelis kontaktas su receptoriais nalorphine daro jį stimuliuojantį poveikį, kuris nesugeba slopinti kvėpavimo centro veiklos, t. turi dalinį agonistinį poveikį opioidų receptoriams, tuo pat metu blokuoja jį morfinui, taip sukeldamas antagonistinį poveikį morfinui. Nalorfino trūkumas yra tas, kad perdozavus, sustiprėja jo stimuliuojantis poveikis receptoriams ir slopinamas kvėpavimo centras, taip pat morfinas.
Tais atvejais, kai vyrauja blokuojantis vaistų farmakologinis poveikis, jie sako, kad vaistas yra antagonistas, turintis savo ar vidinį aktyvumą.
Savo antagonistų veikimo mechanizmo ypatumus galima apsvarstyti acebutalolio β 1 -adrenoblokatoriaus pavyzdžiu. Acebutalolis priklauso β 1 -adrenerginių blokatorių grupei, turintiems savo vidinį simpatomimetinį aktyvumą, t. sujungia β 1 \u200b\u200badrenoreceptorių blokatorių (lizinio) ir stimuliatoriaus (mimetinio) savybes.
Acebutalolio veikimo mechanizmas atsiranda dėl jo cheminės struktūros ypatumų, dėl kurių vaistas, viena vertus, blokuoja miokardo β 1 -adrenoreceptorius, kita vertus, stimuliuoja jų veiklą, taip imituodamas mediatorių - katecholaminų fiziologinį poveikį (žr. T. 1. p. 170).
Visus vaistus pagal jų veikimo ypatumus galima suskirstyti į dvi dideles grupes - turinčius specifinį ir nespecifinį poveikį.
Vaistai, turintys nespecifinį poveikį, apima vaistus, kurie turi platų farmakologinį aktyvumą ir įvairius poveikio taikymo taškus. Šiai grupei priklauso vitaminai, biogeniniai stimuliatoriai, antioksidantai, adaptogenai ir kt.
Vaistai, turintys specifinį poveikį, apima vaistus, pasižyminčius atitinkamų receptorių agonistų ar antagonistų savybėmis, t. jų veikimo mechanizmo pagrindas yra gebėjimas sąveikauti su specifiniais jiems receptoriais.
Tačiau reikia pažymėti, kad vaistų, kurie daro specifinį poveikį bet kuriems receptoriams, farmakologinis poveikis ne visada bus labai specializuotas, pavyzdžiui, mažinant širdies ritmą (HR) ar slopinant skrandžio sulčių sekreciją. Bendras daugelio vaistų farmakologinis poveikis priklausys nuo to, kiek ir kuriuose organuose ir audiniuose yra lokalizuoti šio vaisto receptoriai.
Pavyzdžiui, β-adrenerginiai receptoriai yra širdies raumenyje, kraujagyslių sienoje, bronchuose, gimdoje, riebaliniame audinyje, griaučių raumenyse ir kt. Dėl to vaistai, kurie stimuliuoja β-adrenerginius receptorius, tam tikru mastu sukels širdies susitraukimų stiprumo ir dažnio padidėjimą, kraujagyslių išsiplėtimą, bronchus ir apatinį gimdos tonusą, t. Y., Sisteminį poveikį kūnui.
Konkretus vaistų poveikis gali būti selektyvus arba selektyvus ir atitinkamai neselektyvus arba neselektyvus. Vaistų selektyvumą lemia tai, ar jis veikia visus bet kurių receptorių porūšius, ar jų specifinius porūšius.
Pavyzdžiui, neselektyvus β-adrenoblokatoriaus propranololis blokuoja tiek β 1 -adrenoreceptorius, esančius miokardo audinyje, tiek β 2 -adrenoreceptorius, esančius ypač plaučių audinyje, t. Y. Turi neselektyvų (neselektyvų) poveikį abiems β-adrenerginių receptorių porūšiams. . kadangi terapinės dozės selektyvus β 1 -adrenerginių blokatorių atenololis blokuoja tik β 1 miokardo adrenerginius receptorius, t. daro selektyvų (selektyvų) poveikį specifiniam β-adrenerginiam receptoriui.
Išaiškinus receptorių porūšių fiziologinį vaidmenį ir lokalizaciją, galima sukurti labai efektyvius vaistus, selektyviai veikiančius skirtingus receptorių porūšius.
Pavyzdžiui, vaistai, blokuojantys β 1 -adrenerginius receptorius, klinikoje plačiai naudojami koronarinės širdies ligos, arterinės hipertenzijos gydymui, o vaistai, stimuliuojantys β 2 -adrenerginius receptorius, yra pritaikyti bronchinės astmos gydymui. Klinikinėje praktikoje vaistai, blokuojantys H 1 histamino receptorius, skiriami alerginių reakcijų prevencijai ir (arba) slopinimui, tuo tarpu vaistai, blokuojantys H 2 histamino receptorius, yra veiksmingi gydant skrandžio ir dvylikapirštės žarnos opas.
Sukūrus selektyvųjį poveikį turinčius vaistus, labai pagerėjo jų klinikinis vartojimas, nes jų šalutinis poveikis žymiai sumažėja.
Vaistų veikimo mechanizmo pagrindas, kaip taisyklė, yra jų gebėjimas inicijuoti (suaktyvinti) sudėtingus biocheminius ir (arba) biofizinius procesus, kurie galiausiai keičia ir (arba) optimizuoja tikslinės ląstelės funkcinį aktyvumą.Vaistai gali suvokti jų poveikį organams ir (arba) arba tikslines ląsteles: tiesiogine chemine sąveika; fizikinė ir cheminė sąveika su ląstelės membrana; veiksmai specializuotiems fermentams; veiksmai su reguliavimo genais; veiksmai specifiniams receptoriams.
Tiesioginė cheminė vaistų sąveika.
Šis vaistų veikimo mechanizmas yra gana retas ir gali būti realizuotas už ląstelės ribų, pavyzdžiui, skrandžio ar žarnų spindyje. Jos esmė slypi tame, kad vaistai pradeda tiesioginę cheminę reakciją su molekulėmis ir (arba) jonais, kurie kūne susidaro normalios ar patologinės būklės. Tiesioginės cheminės sąveikos pavyzdys yra cheminė skrandžio druskos rūgšties neutralizacijos reakcija, vartojant antacidinius vaistus.
Fizikinė ir cheminė vaistų sąveika su ląstelės membrana.
Viena iš pagrindinių citoplazminės membranos funkcijų yra jonų mainų tarp citoplazmos ir tarpląstelinės aplinkos įgyvendinimas. Transmembraniniai jonų mainai taip pat gali vykti per specialius nuo įtampos priklausomus transmembraninius jonų kanalus - natrio, kalio, kalcio, chloro ir kt. Kai kurie vaistai, pasiekdami ląstelės membraną, sąveikauja su šiais kanalais ir keičia jų funkcinį aktyvumą. Taigi, pavyzdžiui, antiaritminis IA klasės vaisto - chinidino - poveikis grindžiamas jo gebėjimu blokuoti Na + jonų praėjimą per transmembraninius natrio kanalus.
Vaistų poveikis specializuotiems fermentams.
Palyginti nedidelis kiekis vaistų realizuoja savo farmakologinį poveikį keičiant kai kurių specializuotų ląstelių fermentų aktyvumą. Vaistai, didinantys ląstelių fermentų aktyvumą, yra vadinami induktoriaifermentai. Tokį poveikį turi, pavyzdžiui, migdomosios tabletės ir prieštraukulinis vaistas fenobarbitalis, kuris žymiai padidina mikrosominių kepenų fermentų aktyvumą. Šio fenobarbitalio ir susijusių vaistų biologinė reikšmė bus aptariama toliau.
Vadinami vaistai, slopinantys specializuotų fermentų veiklą inhibitoriaifermentai. Taigi, pavyzdžiui, antidepresantas iš monoaminooksidazės inhibitorių (MAO) grupės, vaistas pirlindolis realizuoja savo antidepresantą, slopindamas MAO fermento aktyvumą centrinėje nervų sistemoje. Gebėjimas slopinti fermento acetilcholinesterazės aktyvumą yra anticholinesterazės vaistų, pavyzdžiui, fizostigmino, farmakologinio aktyvumo pagrindas. Yra žinoma, kad fiziologinėmis sąlygomis acetilcholinesterazė inaktyvuoja (sunaikina) acetilcholiną - neuromediatorių, perduodantį sužadinimą parasimpatinės nervų sistemos sinapsėse. Fizostigminas, slopindamas acetilcholinesterazės aktyvumą, skatina kaupimąsi neurotransmiterio acetilcholino parasimpatinės sistemos sinapsėse, dėl to padidėja parasimpatinės nervų sistemos tonusas, pasireiškiantis sisteminiu lygmeniu vystantis bradikardijai, didėjant virškinimo traktui, mažinant kraujo spaudimą (BP). mokinys ir kt. Vaistai gali grįžtamai ir negrįžtamai sąveikauti su fermentais. Pvz., Vaistas enalaprilis grįžtamai slopina angiotenziną konvertuojančio fermento aktyvumą, kuris visų pirma reiškia kraujospūdžio sumažėjimą, o fosforo organinėms fosforams toksiškos medžiagos negrįžtamai slopina acetilcholinesterazės aktyvumą. Vaistų poveikis norminiams genams.Šiuo metu mokslininkai bando sukurti vaistus, kurie realizuotų savo farmakologinį poveikį tiesiogiai veikdami reguliuojančių genų fiziologinį aktyvumą. Ši tendencija atrodo ypač viltinga po to, kai 2000 m. Buvo iššifruota žmogaus genomo struktūra. Manoma, kad selektyvus reguliavimo genų funkcijos normalizavimas veikiant narkotikams leis pasiekti sėkmės gydant daugelį, įskaitant anksčiau nepagydomas, ligas. Vaistų poveikis receptoriams.Prieš pereinant prie vaistų sąveikos su receptoriais ypatumų, būtina išsiaiškinti, ką turime omenyje terminą „receptorius“ (iš lat. recipio - imk, imk). Nuo fiziologijos eigos žinoma, kad terminas „receptorius“ reiškia labai specializuotas formacijas, gebančias suvokti, transformuoti ir perduoti išorinio signalo energiją nervų sistemai. Šie receptoriai yra vadinami juslinis(iš lat. sensus - jausmas, pojūtis, suvokimas). Jutimo receptoriai apima klausos, regos, uoslės, skonio, lytėjimo organų receptorius. Šių organų jutimo receptoriai priklauso vadinamiesiems eksteroreceptoriams. Jei jutimo organų, reaguojančių į išorinius dirginimo dirgiklius, buvimas buvo žinomas nuo senų senovės, tada jutiminių receptorių buvimas kūne buvo abejotinas iki XIX amžiaus vidurio. Pirmą kartą rusų fiziologas I.F. Zionas, kuris 1866 m. Parodė triušių eksperimento metu sumažėjusį kraujospūdį dėl aortos sudirginimo. Šis atradimas paskatino ieškoti ir tirti receptorius, esančius kūno viduje, ir patys šie receptoriai buvo vadinami interreceptoriai. KamxX amžiaus pradžia buvo atskleistas pakankamas jutiminių interoreceptorių skaičius ir įrodytas jų svarbus vaidmuo reguliuojant fiziologines kūno funkcijas. 1905 m. J. Langley įrodė, kad vaistui tepant ant ląstelės membranos, atsiranda farmakologinis poveikis, jei jis taikomas tik tam tikroje jo vietoje. Be to, ši svetainė sudaro tik nedidelę viso ląstelės paviršiaus ploto dalį. Šis pastebėjimas leido J. Langley daryti išvadą, kad ląstelių membranoje yra specializuotų receptorių vietų, sąveikaujančių su vaistais. Tačiau prioritetas kuriant vaistų veikimo receptorių teoriją priklauso vokiečių fiziologui P. Ehrlichui, kuris 1906 m. Pirmą kartą įvedė terminą „receptoriai“ ir suformulavo postulatą „vaistas neveikia, jei jis nėra pritvirtintas prie ląstelės membranos“. Remiantis P. Ehrlicho teorija, vaisto molekulė turi dvi funkciškai aktyvias grupes, iš kurių viena užtikrina jos fiksaciją ant ląstelės paviršiaus narkotiko receptoriaus srityje, o antroji funkcinė grupė sąveikauja su receptoriais ir sužadina sudėtingą biocheminių reakcijų grandinę, keičiančią jo (ląstelės) fiziologinį aktyvumą. . Taigi, jau XX amžiaus pradžioje. paaiškėjo, kad yra bent dvi interreceptorių klasės: jutimo receptoriaiperduodama informacija apie vidaus organų ir kūno audinių būklę centrinėje nervų sistemoje; ląstelių receptoriaikurie sąveikauja su vaistais, kurie keičia tikslinių ląstelių funkcinį aktyvumą.<>Iškart reikia pastebėti, kad ateityje vadovėlio tekste, siekiant išvengti painiavos terminologijoje, vaistų ir biologiškai aktyvių medžiagų receptoriuose, t. ląstelinis arba cytoreceptorybus žymimi terminu „receptoriai“, tuo tarpu sensoriniai interoreceptoriai bus žymimi terminu, apibūdinančiu jų funkcinį aktyvumą, pavyzdžiui, „baroreceptoriai“, „skausmo receptoriai“ ir kt. P. Ehrlicho atradimas vaistų receptorių ląstelių membranoje buvo atskaitos taškas plėtojant farmakologinį mokslą, ypač farmakodinamiką, kurio vienas iš pagrindinių uždavinių yra ištirti vaistų receptorius veikiančius mechanizmus. Šiuo metu yra atskleista daugybė ląstelių receptorių, tam tikrų biologiškai aktyvių junginių sąveikos su jais ypatumai, kurie leido, viena vertus, suprasti žinomų vaistų veikimo mechanizmą, kita vertus, buvo pagrindas sukurti naujus labai efektyvius vaistus. Natūralu, kad sunku įsivaizduoti, kad evoliucijos metu žmogaus kūne susiformavo įvairių sintetinių (chemiškai gautų) vaistų receptoriai, juo labiau, kad didžioji dauguma šiuolaikinėje farmacijos rinkoje pateikiamų vaistų buvo susintetinta per pastaruosius 50 ar mažiau metų. Įrodyta, kad ląstelės receptorinis aparatas yra labai senas funkcinis-struktūrinis darinys. Taigi, a- ir b-adrenoreceptoriai (receptoriai, kurių sąveika turi norepinefrino ir adrenalino įtaką ląstelės funkciniam aktyvumui) randami ne tik gyvūnų ląstelėse, bet ir augalų ląstelių membranose, pavyzdžiui, augalo nitrozos ląstelėse, kur a- ir b- adrenoreceptoriai reguliuoja protoplazmos (ląstelių turinio) judėjimą. Tuomet kokie yra P. Ehrlicho atrastų vaistų receptoriai ir kodėl jie su jais sąveikauja? Šiuo metu nekyla abejonių, kad vadinamieji vaistų receptoriai iš tikrųjų yra endogeninių (organizme gaminamų) biologiškai aktyvių medžiagų, dalyvaujančių vidaus organų ir kūno audinių funkcinio aktyvumo reguliavime, receptoriai. Tokie biologiškai aktyvūs junginiai apima medžiagas, kurios nervinio signalo perdavimo metu išsiskiria iš nervų galūnių, taip pat hormonus, vitaminus, amino rūgštis ir kt. Kiekvienai endogeninei biologiškai aktyviajai medžiagai yra nustatyti griežtai specifiniai jos receptoriai. Taigi, pavyzdžiui, organizme gaminama biologiškai aktyvi medžiaga adrenalinas gali suaktyvinti griežtai specifinę a - ir b-adrenerginius receptorius, ir gliukokortikosteroidai - antinksčių žievės hormonai - sąveikauja tik su griežtai jiems būdingais gliukokortikosteroidų receptoriais. Sintetiniai vaistai, kurie savo poveikį realizuoja sąveikaudami su ląstelės receptorių aparatais, savo chemine struktūra yra daugiau ar mažiau panašūs į endogeninius biologiškai aktyvius junginius, kurie sąveikauja su panašiais receptoriais. Pavyzdžiui, sintetiniai vazokonstriktoriai (sukeliantys kraujagyslių susiaurėjimą) narkotikai fenilefrinas yra panašūs į endogeninę biologiškai aktyvią medžiagą norepinefriną, todėl, kaip ir norepinefrinas, turi savybę stimuliuoti a-adrenoreceptorius. Kartais dėl savo cheminės struktūros ypatumų vaistai gali sąveikauti ne su pačiu receptoriumi, o su gretima ląstelės membranos dalimi. Kadangi šiuo atveju vaistas sąveikauja ne su pačiu receptoriumi, o su gretima ląstelės membranos dalimi, jie kalba ne apie jaudinantį ar blokuojantį poveikį receptoriui, bet viskasturtingas(iš graikų k alos - kitas, kitoks) efektas arba poveikis. Dėl to gali pasikeisti ir membranos, esančios greta receptoriaus, struktūra, ir atskiri paties receptoriaus komponentai, o tai gali pakeisti receptoriaus jautrumą jam specifiškai biologiškai aktyviajai medžiagai. Tais atvejais, kai padidėja receptoriaus jautrumas biologiškai aktyviajai medžiagai, kalbėkite apie sensibilizacija(iš lat. sensus - jausmas) arba apie sensibilizacija(iš lat. sensibilis - jautrumas) receptorių, ir tais atvejais, kai sumažėja receptoriaus jautrumas, kalbėti apie desensibilizacijareceptorius. Allosterinio efekto ypatumas slypi tame, kad vaistai, turintys tokį veikimo mechanizmą, tiesiogiai nedaro įtakos nervinio impulso perdavimui, bet modifikuoja jį norima linkme. Pavyzdžiui, anksiolitikų (vaistų nuo nerimo; sinonimai: trankvilizatoriai), kurių cheminė struktūra yra benzodiazepino dariniai, veikimo mechanizmas yra pagrįstas postsinapsinių benzodiazepinų receptorių allosterinio sužadinimo reiškiniu. Pastarųjų sužadinimas savo ruožtu skatina slopinančių gama-aminosviesto rūgšties postsinapsinių receptorių (GABA receptorių) aktyvaciją, kuris kliniškai pasireiškia pašalinant neurozinių ligų simptomus, tokius kaip nerimas, nerimas, baimė ir kt. Receptoriai, sąveikaujantys su jais, biologiškai aktyvi medžiaga ar vaistas bet kokiu būdu keičia tikslinės ląstelės funkcinę būklę, yra vadinami konkretus. Be specifinių receptorių, vadinamųjų nespecifinisnarkotikų receptoriams. Specializuotoje medicinos literatūroje šie receptoriai dar vadinami vaistų „praradimo vieta“. Kreipdamiesi į tokius receptorius, vaistai neturi jokio biologinio poveikio, tačiau patys tampa biologiškai neaktyvūs. Šio tipo receptorių pavyzdys gali būti receptoriai, esantys ant plazmos baltymų, ypač ant vandenyje tirpių baltymų - albumino. Receptorių struktūra yra gana sudėtinga, tačiau dauguma jų yra baltymų makromolekulės arba glikoproteinai, kurie taip pat gali apimti jonus, lipidus, nukleorūgštis ir kt. Receptorius t.y. ją sudaranti baltymo makromolekulė pasižymi specifiniu, kiekvienam receptoriui būdingu, jo cheminių grupių erdviniu išdėstymu. Baltymo makromolekulės, sudarančios receptorius, gali būti integruotos (panardintos) į citoplazminės membranos lipidų dvigubą sluoksnį arba lokalizuotos ląstelės viduje. Pagrindinė ląstelės receptoriaus funkcija yra „atpažinti“ cheminį signalą, perduodamą į jį per endogeninę biologiškai aktyvią medžiagą ir (arba) vaistus, ir paversti jį atitinkamu biocheminiu ir (arba) biofiziniu ląstelių atsakymu. Anksčiau buvo manoma, kad vaistai ar endogeninės biologiškai aktyvios medžiagos sąveikauja su „rakto ir užrakto“ tipo receptoriais, t. receptorius turi tokią struktūrą, kuri leidžia vaistui surasti „savo“ receptorių, prisijungti prie jo ir, tarsi, „įjungti“ ir „išjungti“. Tačiau tobulėjant medicinos mokslui tapo akivaizdu, kad tai ne visai tiesa. Šiuo metu tarpląstelinių signalų virsmo ląstelėse, reguliuojančių ląstelių funkciją, molekuliniai procesai jau yra gana gerai ištirti. mechanizmai, sukeliantys endogeninių biologiškai aktyvių medžiagų ar vaistų sąveiką su receptoriais. Sąveikaujant su endogeninės biologiškai aktyvios medžiagos ir (arba) panašaus aktyvaus vaisto receptoriais, jis atsiranda konformacija- baltymo makromolekulės formos erdvinis pokytis, kuris yra įvairių tarpląstelinių procesų, lemiančių tikslinės ląstelės atsaką į tarpininką ir (arba) vaistą, sukėlėjas. Pavyzdžiui, bronchų lygiųjų raumenų b 2 -adrenoreceptorių aktyvinimas veikiant b 2 -adrenostimuliatoriaus fenoteroliui padidina fermento adenilato ciklazės aktyvumą, kuris prisideda prie ciklinio adenozino monofosfato (cAMP) kaupimosi ląstelėje ir dėl to ląstelių atsipalaidavimo. Apskritai, ląsteliniai receptoriai gali būti laikomi griežtai specializuotais ląstelių „jutimo organais“, per kuriuos jie suvokia „informaciją“, kilusią, pavyzdžiui, iš centrinės nervų sistemos ir (arba) endokrininės sistemos. Nepaisant svarbaus receptoriaus aparato vaidmens, receptoriai užima tik nereikšmingą ląstelės membranos dalį. Pvz., Ląstelės M cholinerginių receptorių aparatas užima ne daugiau kaip 1/6 000 jo paviršiaus ploto. Narkotikų sąveikos su receptoriumi savybių tyrimas, viena vertus, leidžia suprasti jo veikimo molekulinio mechanizmo pagrindus, kita vertus, suteikia informacijos apie tai, kokie turėtų būti pokyčiai vaistų struktūroje, siekiant sustiprinti jo gebėjimą sąveikauti su šiuo receptoriumi, t. leidžia tikslingai sintetinti naujus labai efektyvius vaistus. Fiziologinėmis sąlygomis skirtingi ląstelių receptoriai neveikia savarankiškai, bet yra nuolat sąveikaujantys tarpusavyje, taip reguliuodami specifinį ląstelės aktyvumą. Pavyzdžiui, širdies b-adrenerginių receptorių aktyvinimas endogeniniu norepinefrinu ypač sukelia širdies susitraukimų skaičiaus padidėjimą, o širdies M-holinerginių receptorių aktyvacija endogeniniu acetilcholinu, priešingai, sukelia širdies susitraukimų skaičiaus sumažėjimą. Didelis indėlis į vaistų veikimo receptorių mechanizmo supratimą buvo padarytas atradus pre- ir postsinapsinius receptorius. Sinaps(iš graikų k konspektas - jungtis) yra specializuota kontaktinė zona tarp nervų ląstelių ar kitų sužadinamų kūno struktūrų, užtikrinanti gaunamos informacijos perdavimą ir jos informacinės reikšmės išsaugojimą. Sinapsių struktūros ir funkcinio vaidmens tyrimas buvo pradėtas XIX amžiaus pabaigoje. po to, kai ispanų histologas S. Ramon-i-Cajal (S. Ramonas prie Cajal) pasiūlė, kad centrinėje nervų sistemoje būtų specializuota perdavimo sistema. Sinapsės savo pavadinimą įgijo 1897 m., Kai anglų fiziologas Ch. Sherringtonas pasiūlė šį terminą nurodyti kontakto tarp nervų ląstelių plotą. Šiuo metu yra trijų rūšių sinapsės: 1) „elektrinės“ sinapsės, kurių metu informacija perduodama perduodant elektrinį signalą iš priešsinapsinės membranos. Šis sinapsių tipas vadinamas efaps(iš graikų k efapsis - sandarus kontaktas); 2) „cheminės“ sinapsės, kurių metu informacija perduodama per specialias biologiškai aktyvias medžiagas - neurotransmiteriai(iš graikų k neuronas - nervas ir latis. tarpininkas - tarpininkas; sinonimai: tarpininkas); 3) „mišrios“ sinapsės, kurių metu informacija perduodama tiek chemiškai, tiek elektra. Daugelio vaistų, turinčių įtakos sinapsių funkcijoms, farmakologinis poveikis yra realizuojamas jų poveikiu tam tikram signalo perdavimo etapui cheminėse sinapsėse, t. antros rūšies sinapsėse. Paprastai cheminės sinapsės skirstomos pagal neurotransmiterius, perduodančius juose esančius nervinius impulsus, taip: sinapsės, kuriose acetilcholinas veikia kaip tarpininkas, vadinamos cholinerginėmis; sinapsės, kuriose adrenalinas ir norepinefrinas veikia kaip tarpininkas, vadinamos adrenerginėmis; sinapsės, kuriose ATP ir adenozinas veikia kaip tarpininkas, vadinamos purinerginėmis; sinapsės, kuriose gama-aminosviesto rūgštis veikia kaip tarpininkė, vadinamos GABA-erginėmis ir kt. Šiuo metu sinapsių struktūra yra gerai suprantama. Sinapsę sudaro presinapsinis nervinės ląstelės (aksono galo) procesas ir „signalą“ gaunantis aparatas, esantis efektorinės („vykdomosios“) ląstelės membranoje.
Farmakodinamika - farmakologinis poveikis, veikimo mechanizmai, veikimo lokalizavimas, vaistų veikimo tipai.
Farmakologinis poveikis vaistinė medžiaga - organų, kūno sistemų veiklos pokyčiai, kuriuos sukelia ši medžiaga (pavyzdžiui, padidėjęs širdies susitraukimas, sumažėjęs kraujospūdis, stimuliuojantis protinę veiklą, pašalinantis baimę ir įtampą ir kt.).
Kiekviena medžiaga sukelia jai būdingą farmakologinį poveikį. Kiekvienu atveju naudojamas tik tam tikras vaistinės medžiagos poveikis, kuris apibūdinamas kaip pagrindinis padarinius. Likęs (nenaudotas, nepageidaujamas) farmakologinis poveikis vadinamas įkaitas.
Veikimo mechanizmai vaistai - būdai, kuriais medžiagos sukelia farmakologinį poveikį, yra labai įvairūs. Pagrindiniai veiksmų mechanizmų variantai yra šie:
- specifiniai receptoriai;
- fermentai;
- jonų kanalai;
- transporto sistemos.
Dauguma narkotikų veikia specifiniai receptoriai. Šiuos receptorius dažniausiai atspindi funkciškai aktyvios baltymų molekulės, kurių sąveika sukelia biochemines reakcijas, sukeliančias farmakologinį poveikį.
Yra specifiniai receptoriai, susiję su ląstelių membranomis (membrana), ir tarpląsteliniai receptoriai (citoplazminė, branduolinė).
Membranos receptoriai (citoplazminės membranos receptoriai) skirstomi į:
- receptoriai, tiesiogiai sujungti su jonų kanalais;
- receptoriai, tiesiogiai konjuguoti su fermentais;
- receptoriai, sąveikaujantys su G baltymais.
Kam receptoriai, tiesiogiai sujungti su jonų kanalaisPirmiausia apima include-cholinerginius receptorius ir GABA A receptorius.
Kai stimuliuojami Ν cholinerginiai receptoriai (jautrūs nikotinui cholinerginiai receptoriai), tiesiogiai su jais sujungti natrio kanalai atsidaro. Stimuliuojant Ν-cholinerginius receptorius atsiranda Na + kanalų atradimas, Na + jonų patekimas į ląstelę, ląstelės membranos depoliarizacija ir jaudinantis poveikis.
GABA A receptoriai yra tiesiogiai sujungti su chloro kanalais. GABA A receptorių stimuliacija lemia Cl kanalų atradimą, Cl jonų patekimą, ląstelės membranos hiperpolarizaciją ir slopinamąjį poveikį.
Kam receptorius, tiesiogiai susijusius su fermentaisTarp jų, visų pirma, yra insulino receptorių, tiesiogiai konjuguotų su tirozino kinaze.
G baltymų receptoriai - M-cholinerginiai receptoriai (raumenims jautrūs cholinerginiai receptoriai), adrenerginiai receptoriai, dopamino receptoriai, opioidiniai receptoriai ir kt.
G baltymai, t. Y. GTP surišantys baltymai, yra lokalizuoti ląstelės membranoje ir susideda iš α-, β- ir γ-subvienetų. Kai vaistas sąveikauja su receptoriais, G-baltymo α-subvienetas jungiasi su GTP (GTP) ir veikia fermentus arba jonų kanalus.
Vienas receptorius sąveikauja su keliais G baltymais, o kiekvienas G baltymo α-subvienetų ir GTP kompleksas veikia keletą fermento molekulių arba kelis jonų kanalus. Taigi įgyvendinamas stiprintuvo (stiprintuvo) mechanizmas: suaktyvinus vieną receptorių, keičiasi daugelio fermento molekulių ar daugelio jonų kanalų aktyvumas.
Vienas pirmųjų buvo rasta G baltymų, susijusių su širdies β 1 adrenoreceptoriais. Suaktyvinus simpatinę širdies inerciją, β 1 -adrenoreceptoriai sužadinami; adenilato ciklazė aktyvuojama per G-baltymus; Iš ATP susidaro cAMP, aktyvuojama baltymų kinazė, veikianti Ca 2+ kanalus fosforilinant ir atidarant.
Padidėjęs Ca 2+ jonų įnešimas į sinoatrialinio mazgo ląsteles pagreitina ketvirtąją veiksmo potencialo fazę, padidėja generuojamų impulsų dažnis - širdies susitraukimai tampa greitesni.
Aptikus Ca 2+ kanalus darbinio miokardo skaidulose, padidėja Ca 2+ koncentracija citoplazmoje (Ca 2+ įvedimas skatina Ca 2+ išsiskyrimą iš sarkoplazminio retikulumo). Ca 2+ jonai jungiasi su troponinu C (troponino-tropomiozino komponentu); Taigi slopinamas troponino-tropomiozino poveikis aktino ir miozino sąveikai mažėja - sustiprėja širdies susitraukimai (10 pav.).
Fig. 10. Širdies susitraukimų didinimo ir stiprinimo mechanizmas stimuliuojant β 1 -adrenoreceptorius. AC - adenilato ciklazė; PC - baltymų kinazė; CA - sinoatrialinis mazgas; Ttm - troponinas-tropomiozinas.
Suaktyvinus parasimpatinę širdies (vagos nervų) inervaciją, sužadinami M2-cholinerginiai receptoriai, o per G-baltymus slopinamas adenilato ciklazė - silpnėja ir susilpnėja širdies susitraukimai (silpnėja daugiausia prieširdžių susitraukimai, nes skilvelių parasimpatinė inervacija yra gana prasta).
Taigi G-baltymai gali stimuliuoti ir slopinti adenilato ciklazę. Stimuliuojantys G baltymai buvo žymimi kaip G s baltymai (stimuliuoja), o slopinantys G baltymai - G baltymai (slopina) (11 pav.).
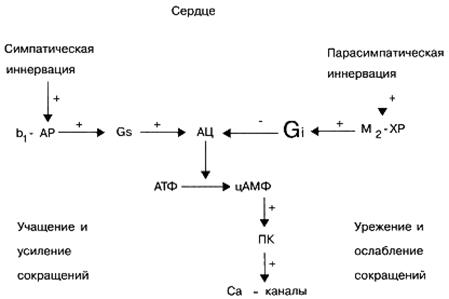
Fig. 11. Širdies susitraukimų dažnio ir stiprumo pokyčių mechanizmas stimuliuojant simpatinę ir parasimpatinę inervaciją.
Choleros toksinas suaktyvina C baltymus (tai sąlygoja adenilato ciklazės aktyvaciją, o choleroje pasireiškia skysčio sekrecija per žarnyno epitelį).
Pertussis toksinas aktyvina G i baltymus.
Kai M 1 -cholinerginiai receptoriai, M3 -cholinerginiai receptoriai, α1 -adrenoreceptoriai sužadinami per Gq baltymus, suaktyvėja fosfolipazė C, kuri prisideda prie inozitol-1,4,5-trifosfato ir diacilglicerolio susidarymo iš fosfatidilinozitol-4,5-difosfato. .
Inozitol-1,4,5-trifosfatas veikia jautrius sarkoplazminės retikulinės membranos receptorius ir stimuliuoja Ca 2+ jonų išsiskyrimą iš sarkoplazminio retikulumo (12 pav.). Stimuliuojant kraujagyslių αι-adrenoreceptorius, tai sąlygoja kraujagyslių lygiųjų raumenų susitraukimą ir kraujagyslių susiaurėjimą (13 pav.).

Fig. 12. Fosfolipazės C poveikis citoplazmos C a2 + lygiui.
Receptorių jautrumas agonistams ir receptorių skaičius nuolat kinta. Taigi, stimuliavus β 1 -adrenoreceptorius β 1 -adrenoreceptorių agonistu, fosforilinamas specialiu receptorių kinaze, jis jungiasi su β-arrestino baltymu ir šiame komplekse praranda gebėjimą sąveikauti su G-baltymais (receptorių desensibilizacija). Β 1 -adrenoreceptorių kompleksą su β-arrestinu ląstelė absorbuoja per endocitozę (receptorių internalizaciją) ir jį sugauna endosomos ir lizosomos. Endosomose β 1 -restino molekulės yra atskirtos nuo receptorių, kurie vėl integruojasi į ląstelės membraną; receptorių jautrumas agonistams yra atkuriamas (receptorių jautrumas). Lizosomose pažeidžiamos receptorių molekulės (sumažėjęs reguliavimas) (14 pav.).

Fig. 13. Kraujagyslių lygiųjų raumenų susitraukimo mechanizmas simpatinės inervacijos stimuliavimo metu. FLS - fosfolipazė C; FIF 2 - fosfatidilinozitol-4,5-difosfatas; Jei 3 - inozitol-1,4,5-trifosfatas; SR - sarkoplazminis retikulumas; KLCM - miozino lengvųjų grandinių kinazė.

Fig. 14. β-adrenerginių receptorių jautrumas ir slopinimas.
Kam tarpląsteliniai receptoriai kortikosteroidų ir lytinių hormonų receptoriai yra įtraukti. Visų pirma, gliukokortikoidų receptoriai yra lokalizuoti ląstelių citoplazmoje. Derinant gliukokortikoidus su citoplazmos receptoriais, gliukokortikoidų - receptorių kompleksas prasiskverbia į branduolį ir daro įtaką įvairių genų ekspresijai.
Medžiagų gebėjimą jungtis prie receptorių (medžiagų polinkį jungtis prie receptorių) rodo terminas „ giminystė“. Tų pačių receptorių atžvilgiu skirtingų medžiagų afinitetas gali būti skirtingas. Norėdami apibūdinti giminingumą, naudokite indikatorių pKD yra neigiamas disociacijos konstantos logaritmas, tai yra medžiagos koncentracija, kurioje užimta 50% receptorių.
Vidinė veikla - medžiagų gebėjimas stimuliuoti receptorius; nustatomas pagal farmakologinio poveikio, susijusio su receptorių aktyvacija, dydį.
Normaliomis sąlygomis nėra tiesioginio ryšio tarp giminingumo ir vidinio aktyvumo. Medžiaga gali užimti visus receptorius ir sukelti silpną poveikį, ir atvirkščiai, medžiaga gali užimti 10% receptorių ir sukelti maksimalų šios sistemos poveikį.
Agonistai - giminingumo ir vidinio aktyvumo medžiagos.
Visiški agonistai pasižymi giminingumu ir maksimaliu vidiniu aktyvumu (jie gali sukelti maksimalų efektą tam tikroje sistemoje), net jei jie užima dalį specifinių receptorių.
Daliniai (daliniai) agonistai pasižymi giminingumu ir mažesniu nei maksimaliu vidiniu aktyvumu (jie gali sukelti tik mažesnį nei maksimalų poveikį, net jei jie užima 100% specifinių receptorių).
Antagonistaituri afinitetą, tačiau neturi vidinio aktyvumo ir trukdo visiškai ar iš dalies agonistams veikti (pašalina agonistus nuo ryšio su receptoriais).
Jei antagonisto poveikis pašalinamas padidinus agonisto dozę, toks antagonizmas vadinamas konkurenciniu.
Daliniai agonistai gali būti visiškų agonistų antagonistai. Jei nėra visiško agonisto, dalinis agonistas stimuliuoja receptorius ir sukelia silpną poveikį. Sąveikaudamas su visišku agonistu, dalinis agonistas užima receptorius ir slopina visiško agonisto veikimą. Tokiu atveju susilpnėja visiško agonisto poveikis.
Pavyzdžiui, pindololis, dalinis β-adrenoreceptorių agonistas, sukelia silpną tachikardiją, jei širdyje nėra simpatinės inervacijos poveikio. Tačiau padidėjus simpatinės inervacijos tonui, pindololis veikia kaip tikras β blokatorius ir sukelia bradikardiją. Taip yra dėl to, kad dalinis agonistas pindololis silpnina tarpininko norepinefrino, kuris yra visiškas agonistas, poveikį β-adrenerginiams širdies receptoriams, poveikį.
Antagonistų agonistai - medžiagos, kurios skirtingai veikia tų pačių receptorių potipius: stimuliuoja vienus receptorių potipius, o kitus blokuoja. Pvz., Narkotinis analgetikas nalufinas daro skirtingą poveikį opioidų receptorių potipiams. Nalbufinas stimuliuoja κ receptorius (todėl sumažina jautrumą skausmui), o μ receptoriai blokuoja (todėl yra mažiau pavojingi priklausomybės nuo narkotikų atžvilgiu).
Medžiagų poveikio pavyzdys fermentai gali būti anticholinesterazės agentų, kurie blokuoja acetilcholinesterazę (fermentą, kuris skaido acetilcholiną) ir tokiu būdu sustiprina ir prailgina acetilcholino veikimą.
Žinomos vaistinės medžiagos, kurios stimuliuoja ar blokuoja jonų kanalai ląstelių membranos, tai yra kanalai, selektyviai vedantys Na +, K +, Ca 2+ jonus (natrio, kalio, kalcio kanalus) ir kt. Pavyzdžiui:
Vietiniai anestetikai blokuoja Na + kanalus;
I klasės antiaritminiai vaistai (chinidinas, lidokainas) blokuoja Na + kanalus;
Minoksidilis aktyvina K + kanalus;
Hipoglikeminiai agentai iš sulfonilkarbamido darinių grupės blokuoja nuo ATP priklausomus K + kanalus;
Verapamilis, nifedipinas blokuoja Ca 2+ kanalus.
Medžiagų poveikio pavyzdys transporto sistemos gali būti veiksmas:
Rezerpinas (blokuoja vezikulinį dopamino ir norepinefrino įsisavinimą);
Širdies glikozidai (slopina Na + / K + -ATPazę);
Tricikliai antidepresantai (blokuoja norepinefrino ir serotonino pasipriešinimą neuronams);
Protonų siurblio blokatoriai (omeprazolas ir kt.).
Galimi ir kiti veikimo mechanizmai. Pavyzdžiui, diuretikas manitolis padidina diurezę padidindamas osmosinį slėgį inkstų kanalėliuose. Antiaterosklerozinis agentas - kolestipolis - suriša (secesteruoja) tulžies rūgštis, trukdo jų pasisavinimui žarnyne, todėl kepenyse suaktyvėja tulžies rūgščių susidarymas iš cholesterolio ir sumažėja hepatocitų cholesterolio lygis.
Įvairių vaistų veikimo mechanizmai buvo ištirti skirtingai. Studijuodami jas, idėjos apie veikimo mechanizmus gali ne tik tapti sudėtingesnės, bet ir žymiai pasikeisti.
Sąvoka „ veiksmo lokalizavimas"Reiškia tam tikrų vaistinių medžiagų pagrindinę veikimo vietą (-as). Pavyzdžiui, širdies glikozidai daugiausia veikia širdį.
Į koncepciją „ veiksmų tipai»Įtraukite vietinius ir bendruosius (rezorbcinius) veiksmus, refleksinius veiksmus, pagrindinius ir šalutinius poveikius, tiesioginius ir netiesioginius veiksmus.
Vietinio veiksmo pavyzdys gali būti vietinių anestetikų veikimas.
Daugelis vaistų turi bendrąjį (rezorbcinį) poveikį, kuris paprastai išsivysto absorbavus (rezorbciją) medžiagą kraujyje ir pasiskirstant organizme.
Tiek vietinio, tiek rezorbcinio poveikio metu medžiagos gali sužadinti įvairius jautrius receptorius ir sukelti refleksines reakcijas.
Pagrindinis vaistinės medžiagos poveikis yra jos poveikis, kuris naudojamas kiekvienu atveju. Visi kiti poveikiai vertinami kaip šalutinio poveikio pasireiškimai.
Vaistinės medžiagos gali turėti tiesioginį poveikį tam tikriems organams. Be to, narkotikų poveikis gali būti netiesioginis. Pavyzdžiui, širdies glikozidai daro tiesioginį poveikį širdžiai, tačiau, pagerindamos širdies darbą, šios medžiagos padidina kraujo tiekimą ir kitų organų funkcijas (netiesioginis poveikis).