اکثریت قریب به اتفاق داروها داشتن اثر درمانی با تغییر فعالیت سیستمهای فیزیولوژیکی سلولهایی که در طول تکامل در بدن تولید می شوند. تحت تأثیر یک ماده دارویی در بدن ، به عنوان یک قاعده ، نوع جدیدی از فعالیت سلولی بوجود نمی آید ، فقط نرخ انواع مختلف فرایندهای طبیعی تغییر می کند. مهار یا تحریک فرآیندهای فیزیولوژیکی منجر به کاهش یا تقویت عملکردهای مربوط به بافتهای بدن می شود.
داروها می توانند بر روی گیرنده ها ، آنزیم ها ، غشاهای سلولی خاص عمل کنند یا به طور مستقیم با مواد سلولی ارتباط برقرار کنند. جزئیات مکانیسم های عمل مواد دارویی در دوره ای از فارماکولوژی عمومی یا تجربی مورد مطالعه قرار گرفته است. در زیر فقط چند نمونه از مکانیسم های اصلی فعالیت داروها را ارائه می دهیم.
تأثیر بر گیرنده های خاص. گیرنده ها ساختارهای ماکرومولکولی هستند که از نظر انتخابی نسبت به برخی ترکیبات شیمیایی حساس هستند. تعامل مواد شیمیایی با گیرنده منجر به بروز تغییرات بیوشیمیایی و فیزیولوژیکی در بدن می شود ، که در یک اثر بالینی خاص بیان شده است.
داروهایی که مستقیماً فعالیت عملکردی گیرنده ها را تحریک می کنند یا افزایش می دهند ، آگونیست نامیده می شوند و موادی که مانع از عملکرد آگونیست های خاص می شوند ، آنتاگونیست نامیده می شوند. آنتاگونیسم می تواند رقابتی و غیر رقابتی باشد. در حالت اول ، ماده مخدر برای اتصال سایتها در گیرنده های خاص با یک تنظیم کننده طبیعی (واسطه) رقابت می کند. محاصره گیرنده ناشی از آنتاگونیست رقابتی با دوزهای زیادی از آگونیست یا یک واسطه طبیعی از بین می رود.
انواع گیرنده ها با حساسیت به واسطه های طبیعی و آنتاگونیست های آنها تقسیم می شوند. به عنوان مثال ، گیرنده های حساس به استیل کولین به کولینرژیک گفته می شوند ، گیرنده های حساس به آدرنالین را آدرنرژیک می نامند. از نظر حساسیت به موسکارین و نیکوتین ، گیرنده های کولینرژیک به حساس به موسکارینی (گیرنده های مایع کولینرژیک) و حساس به نیکوتین (گیرنده های N-cholinergic) تقسیم می شوند. گیرنده های H- کولینرژیک ناهمگن هستند. مشخص شده است که تفاوت آنها در حساسیت نسبت به آنها نهفته است مواد مختلف. گیرنده های n-کولینرژیک در گانگلیون سیستم عصبی خودمختار ، و گیرنده های n-کولینرژیک عضله مری وجود دارد. انواع مختلفی از گیرنده های آدرنرژیک شناخته شده است ، با حروف یونانی α1 ، α2 ، β1 ، β2.
هیستامین H1 و H2 ، دوپامین ، سروتونین ، مواد افیونی و سایر گیرنده ها نیز جدا شده اند.
تأثیر فعالیت آنزیم. برخی داروها فعالیت آنزیمهای خاص را افزایش داده و یا مهار می کنند. به عنوان مثال ، فیزوستیگمین و نئوستیگمین فعالیت کولین استراز را کاهش می دهند ، که استیل کولین را از بین می برد و اثرات مشخصه برانگیختگی سیستم عصبی پاراسمپاتیک را از بین می برد. مهار کننده های مونوآمین اکسیداز (ایپرازید ، نیالامید) که از تخریب آدرنالین جلوگیری می کنند ، باعث افزایش فعالیت سیستم عصبی سمپاتیک می شوند. فنوباربیتال و زیکسورین با افزایش فعالیت گلوکورونیل ترانسفراز کبد ، سطح بیلی روبین در خون را کاهش می دهد.
اثرات فیزیکی و شیمیایی بر روی غشای سلولی. فعالیت سلولهای سیستم عصبی و عضلانی به جریان یونی بستگی دارد که پتانسیل الکتریکی ترانسمیبرون را تعیین می کند. برخی از داروها انتقال یون را تغییر می دهند.
بنابراین داروهای ضد آریتمی ، ضد تشنج ، داروهای بیهوشی عمومی.
تعامل شیمیایی مستقیم. داروها می توانند به طور مستقیم با مولکول های کوچک یا یون های داخل سلول ارتباط برقرار کنند. به عنوان مثال ، اسید اتیلن دی آمینتراستریک (EDTA) به شدت یون های سرب را به هم متصل می کند. اصل اثر متقابل شیمیایی مستقیم مبتنی بر استفاده از بسیاری از پادزهرها برای مسمومیت شیمیایی است. نمونه دیگر خنثی سازی اسید هیدروکلریک با آنتی اسیدها است.
فارماکودینامیک
این مطالعه مکانیسم عملکرد داروها و همچنین اثرات بیوشیمیایی و فیزیولوژیکی آنها را مورد بررسی قرار می دهد. وظایف وی شامل توصیفی از فعل و انفعالات شیمیایی و فیزیکی بین دارو و سلول هدف و همچنین طیف و شدت کامل اثرات دارویی آن است. آگاهی از الگوهای فارماکودینامیک به شما امکان می دهد داروی مناسب را انتخاب کنید. مطالعات فارماکودینامیکی درک عمیق تری از تنظیم فرآیندهای بیوشیمیایی و فیزیولوژیکی در بدن ارائه می دهد (Katzung B.G.، 1998؛ Lawrence D.R. et al.، 2002).
عمل بیشتر داروها با اتصال آنها به ماکرومولکول های بدن واسطه می شود. تغییر در وضعیت عملکردی این ماکرومولکولها ، به نوبه خود ، باعث ایجاد زنجیره ای از واکنشهای بیوشیمیایی و فیزیولوژیکی می شود که به یک اثر دارویی تبدیل می شوند. ماکرومولکول هایی که مواد شیمیایی با آنها در ارتباط هستند ، گیرنده نامیده می شوند. بنابراین ، هر ماکرومولکول های عملکردی فعال می توانند به عنوان گیرنده داروها باشند. چندین نتیجه مهم از این بیانیه تبعیت می کند. ابتدا با کمک داروها می توانید سرعت هر فرآیند فیزیولوژیکی در بدن را تغییر دهید. ثانیاً ، داروها فقط عملکردهای فیزیولوژیکی طبیعی سلول را تغییر می دهند ، بدون اینکه به آن خواص جدیدی بدهید.
گیرنده ها
بیشتر گیرنده ها پروتئین هستند. این گیرنده های هورمون ها ، فاکتورهای رشد ، واسطه ها ، پروتئین های دخیل در مهمترین واکنش های متابولیکی و نظارتی (دی هیدروفولات ردوکتاز ، استیل کولین استراز) ، پروتئین های حمل و نقل (Na + ، K + -ATPase) ، پروتئین های ساختاری (توبولین) هستند. اجزای سلولی با ماهیت شیمیایی متفاوت مانند اسیدهای نوکلئیک که عوامل ضد تومور با آنها در ارتباط هستند ، همچنین می توانند به عنوان گیرنده عمل کنند.
گیرنده های عوامل نظارتی درون زا - هورمون ها ، واسطه ها و غیره ، اهمیت دارویی دارند. این گیرنده ها به عنوان اهداف بسیاری از داروها خدمت می کنند ، معمولاً به دلیل خاصیت بالای گیرنده ها برای لیگاند های درون زا به طور انتخابی عمل می کنند. داروهایی که پس از اتصال به گیرنده ، اثر فیزیولوژیکی لیگاند درون زا را تولید می کنند ، آگانیست یا محرک نامیده می شوند. داروهایی که این اثر را ایجاد نمی کنند ، اما اتصال لیگاند های درون زا را مهار می کنند ، آنتاگونیست یا مسدود کننده نامیده می شوند. موادی که تأثیر آن کمتر از اثر آگونیستها است ، آگونیستهای جزئی نامیده می شوند. آماده سازی های ایجاد کننده گیرنده در یک ترکیب غیرفعال به عنوان آگونیست های معکوس طبقه بندی می شوند.
وابستگی ساختاری و عملکردی
ساختار شیمیایی دارو نسبتاً سفت و سخت میل آن را به گیرنده ها و فعالیت های داخلی تعیین می کند. تغییر جزئی در ساختار شیمیایی می تواند به طور قابل توجهی بر خواص دارویی تأثیر بگذارد.
سنتز داروهای جدید تا حد زیادی بر این اساس استوار است. از آنجا که اصلاح شیمیایی لزوماً بر تمام خصوصیات دارویی تأثیر نمی گذارد ، می توان اثر بخشی و ایمنی دارو را افزایش داد ، گزینش آن را افزایش داده و ویژگی های فارماکوکینتیک را بهبود بخشید. به عنوان مثال ، بسیاری از آنتاگونیست های هورمون و واسطه مورد استفاده در کلینیک با اصلاح شیمیایی مواد درون زا سنتز می شوند.
نکات کاربرد مواد مخدر
از آنجا که تأثیر دارو توسط گیرنده ها واسطه می گیرد ، نقطه استفاده از دارو نه تنها از ویژگی های توزیع آن ، بلکه با بومی سازی گیرنده ها مشخص می شود و اثرات دارویی به اهمیت عملکردی این گیرنده ها بستگی دارد. اثرات دارویی داروهایی که گیرنده های آنها در بسیاری از ارگان ها و بافت ها معمول است ، متنوع است. اگر این گیرنده ها عملکردی را برای سلول ها انجام دهند ، استفاده از این دارو فقط برای اهداف درمانی دشوار نیست ، بلکه همچنین ناامن است. با این وجود ، چنین داروهایی می توانند از اهمیت بالینی بالایی برخوردار باشند. بنابراین ، گلیکوزیدهای قلبی ، که به طور گسترده در نارسایی قلبی مورد استفاده قرار می گیرد ، انتقال یونها از طریق غشای سلولی را تغییر می دهند ، که فعالیت حیاتی سلول به آن بستگی دارد. آنها دامنه درمانی باریکی دارند و بسیار سمی هستند. مثال دیگر عوامل ضد توموری است. اگر گیرنده هایی که دارو با آنها تداخل دارند فقط در چند نوع سلول متفاوت وجود داشته باشند ، تأثیر آن انتخابی تر است. این داروها ممکن است عوارض جانبی کمتری داشته باشند ، اما اگر گیرنده های آنها عملکرد حیاتی داشته باشند ، ممکن است این داروها سمی باشند. بعضی از سموم بیولوژیکی (سم بوتولینوم و غیره) به همان روش عمل می کنند. علاوه بر این ، حتی اگر اثر دارویی مستقیم انتخابی باشد ، پیامدهای آن ممکن است متنوع تر باشد.
گیرنده های فاکتور تنظیم کننده درون زا
اصطلاح گیرنده به هر مؤلفه ماکرومولکولی سلول که با آن یک دارو وصل می شود ، اطلاق می شود. یکی از مهمترین گیرنده های دارویی پروتئین های سلولی است که به عنوان گیرنده های عوامل نظارتی درون زا - هورمون ها ، فاکتورهای رشد ، واسطه ها - خدمت می کنند. با اتصال به لیگاند درون زا ، گیرنده ها سیگنال را از آن به سلول مورد نظر منتقل می کنند.
از گیرنده ، سیگنال به طور مستقیم یا از طریق مولکول های سیگنالینگ واسطه - مبدل پروتئین - به اهداف سلولی (پروتئین های مؤثر) می رسد. گیرنده ، مبدل پروتئین و پروتئین های مؤثر سیستم گیرنده-مؤثر را تشکیل می دهند. نزدیکترین پروتئین مؤثر در زنجیره انتقال سیگنال معمولاً یک مؤثر ترمینال نیست (مستقیماً بر عملکردهای سلولی تأثیر می گذارد) ، بلکه یک آنزیم یا پروتئین حمل و نقل است که در شکل گیری ، انتقال یا غیرفعال کردن یک واسطه دوم - یونی یا یک مولکول کوچک نقش دارد. واسطه دوم ، به نوبه خود ، اطلاعات را به انواع اهداف درون سلولی منتقل می کند ، و پاسخ همزمان آنها به یک سیگنال از یک گیرنده را تضمین می کند.
گیرنده ها ، پروتئین های تبدیل کننده و پروتئین های مؤثر نه تنها اطلاعات را منتقل می کنند. آنها همچنین از یک سو سیگنال های لیگاند های مختلف را هماهنگ می کنند و از طرف دیگر تمام این سیگنال ها را با فرآیندهای متابولیکی در سلول انجام می دهند.
گیرنده ها به عنوان کاتالیزور ، سیگنال بیولوژیکی را تقویت می کنند. با توجه به این خاصیت مهم ، آنها به عنوان اهداف عالی برای داروها خدمت می کنند. با این حال ، تقویت کننده های سیگنال نه تنها گیرنده هایی با فعالیت آنزیمی هستند ، بلکه همه گیرنده های شناخته شده نیز هستند. در واقع ، هنگامی که یک مولکول لیگاند تنها به یک گیرنده کونژوگه شده به یک کانال یونی متصل می شود ، بسیاری از یون ها از حالت دوم عبور می کنند. همین امر در مورد گیرنده های هورمون استروئیدی نیز صادق است: یک مولکول هورمون باعث رونویسی بسیاری از کپی های mRNA می شود ، که بر اساس آنها مولکول های پروتئینی متعددی ساخته می شوند.
بسته به ساختار و مکانیسم عمل ، گیرنده های مواد فعال بیولوژیکی به چند طبقه تقسیم می شوند. تعداد این کلاس ها اندک است.
گیرنده های آنزیمی
بزرگترین گروه گیرنده هایی که فعالیت آنزیمی دارند گیرنده های غشایی با فعالیت پروتئین کیناز خود هستند. آنها پروتئین های مؤثر واقع در داخل غشای سلولی را فسفریله می کنند. در نتیجه ، عملکرد این پروتئین ها یا تعامل آنها با پروتئین های دیگر تغییر می کند.
کلاس دیگری از گیرنده ها با فعالیت پروتئین کیناز وجود دارد - این گیرنده ها با پروتئین کینازها ترکیب شده اند. آنها فاقد دامنه کاتالیزوری داخل سلولی هستند ، اما هنگام تعامل با آگونیست ، پروتئین کیناز درون سلولی را در سطح داخلی غشا متصل یا فعال می کنند. این گیرنده ها برای عوامل عصبی و گیرنده های تشخیص دهنده آنتی ژن برای لنفوسیت های T و B متشکل از چندین زیر واحد هستند. دومی همچنین با فسفاتیروزین فسفاتها در تعامل است. عملکرد گیرنده های دیگر که دامنه تأثیر داخل سلولی ندارند ممکن است توسط برخی پروتئین های مؤثر دیگر واسطه شود.
گیرنده های دیگر با فعالیت آنزیمی خود ساختار مشابهی دارند. اینها شامل ، به عنوان مثال ، گیرنده هایی با فعالیت فسفاتیروزین فسفاتاز خاص خود هستند: دامنه خارج سلولی آنها در توالی اسید آمینه با مولکولهای چسبندگی مشابه است. برای بسیاری از گیرنده ها با فعالیت فسفاتیروزین فسفاتاز خاص خود ، لیگاندهای درون زا شناخته نمی شوند. اما طبق مطالعات ژنتیکی و بیوشیمیایی انجام شده بر روی انواع مختلف سلولها ، فعالیت آنزیمی این گیرنده ها نقش مهمی دارند. دامنه درون سلولي گيرنده هاي هورمون ناتريورتيك دهليزي ، ساير NUP ها و همچنين گيرنده هاي گوانيلين ، فعاليت گوانلات سيكلاز خاص خود را دارد و cGMP را كه به عنوان واسطه دوم عمل مي كند ، سنتز مي كند. شاید گیرنده های دیگری نیز با فعالیت آنزیمی خاص خود وجود داشته باشند.
گیرنده های همراه کانال یونی
گیرنده های برخی واسطه ها مستقیماً با کانال های یونی در ارتباط هستند و با یک لیگاند در تعامل هستند تا یون های خاصی را از طریق غشای سلولی منتقل کنند (کانال های حساس کننده ، کانال های گیرنده یونیوتروپیک ، گیرنده های یونوتروپیک).
گیرنده های همراه پروتئین G
این یک کلاس گیرنده نسبتاً بزرگی است که از طریق پروتئینهای G (پروتئین هایی که از تعویض گوانین دی فسفات (GDF) برای تری فسفات گوانین (GTP)) استفاده می کنند ، تأثیر متقابل دارد. و پروتئین های لیگاند: آنزیم ها (آدنیلات سیکلاز ، فسفولیپاز C) و کانال های غشایی پتاسیم و کلسیم به عنوان مؤثر عمل می کنند.تعداد زیاد و نقش فیزیولوژیکی مهم گیرنده های همراه با پروتئین های G آنها را بسیار عالی می کند. اهداف من برای داروها: تقریبا نیمی از داروهای تجویز شده توسط پزشکان (به استثنای آنتی بیوتیک ها) روی این گیرنده ها عمل می کنند.
یک سلول می تواند حداکثر 20 گیرنده را بر روی سطح خود داشته باشد که هر یک به طور انتخابی با یک یا چند نوع پروتئین G در تعامل هستند (در انواع مختلف زیر واحد α متفاوت است). زیر واحد α قادر به تعامل با یک یا چند پروتئین مؤثر است ، که به شما امکان می دهد سیگنال های گیرنده های لیگاند های مختلف را با استفاده از یک پروتئین G هماهنگ کنید. از طرف دیگر ، یک گیرنده منفرد می تواند چندین مکانیسم انتقال سیگنال داخل سلولی را فعال کند ، چندین نوع پروتئین G را فعال می کند و از طریق همان زیر واحد α بر روی پروتئین های مختلف مؤثر عمل می کند. چنین سیستم پیچیده واگرایی و همگرایی سیگنالها تنظیم انعطاف پذیر عملکردهای سلولی را فراهم می کند (راس ، 1992).
گیرنده های داخل سلولی
گیرنده های هورمون های استروئیدی و تیروئید ، کلسیتریول و رتینوئیدها پروتئین های محلول اتصال DNA درون سلول هستند که تنظیم رونویسی از ژن های خاصی را کنترل می کنند (Mangelsdorf et al.، 1994). این گیرنده ها متعلق به فوق العاده تنظیم کننده های رونویسی حساس به لیگاند هستند. عملکرد فاکتورهای رونویسی توسط فسفوریلاسیون ، تعامل با پروتئینهای سلولی ، متابولیتها و سایر اجزای نظارتی سلول تنظیم می شود.
سیستم های واسطه دوم
cAMP: سیستم های واسطه ثانویه نیز در ادغام سیگنال های خارجی نقش دارند. اگرچه گیرنده ها و مولکول های سیگنالینگ پروتئین بسیار شناخته شده تر از واسطه های دوم هستند ، در حالت دوم در بسیاری از مسیرهای انتقال سیگنال سلولی نقش دارند. واسطه های دوم مورد مطالعه شامل cAMP ، cGMP ، Ca 2+ ، IF 3 (تری فسفات اینوزیتول) ، DAG (دیاسیل گلیسرول) ، NO. این گروه از ترکیبات ناهمگن به طور مداوم در حال رشد هستند. واسطه های دوم به طور مستقیم (با تغییر متابولیسم یکدیگر) یا غیرمستقیم (با اقدام به همان اهداف درون سلولی) ارتباط برقرار می کنند. عملکرد واسطه های دوم و همچنین تنظیم شکل گیری آنها (یا رهاسازی) ، برش و دفع از سلول ، به راحتی با مثال cAMP در نظر گرفته می شود. این واسطه دوم تحت تأثیر آدنیلات سیتاز با فعال شدن بسیاری از گیرنده های کونژوگه شده با پروتئین های G سنتز می شود. پروتئین G باعث فعال شدن آدنیلات سیکلاز می شود ، پروتئین G باعث مهار می شود.
حداقل 10 ایزوفرم آدنیلات سیکلوتاز خاص در بافت وجود دارد که در مکانیسم های تنظیم فعالیت متفاوت است.
به عنوان یک قاعده ، اردوگاه پروتئین کیناز A (پروتئین کیناز وابسته به اردوگاه) ، گروه کوچکی از پروتئین های مرتبط را فعال می کند. این پروتئین کینازها ، به نوبه خود ، فسفریلات نه تنها اهداف نهایی داخل سلولی (آنزیم ها ، پروتئین های حمل و نقل) ، بلکه پروتئین کینازهای دیگر و سایر پروتئین های نظارتی را نیز به خود اختصاص می دهند. بعنوان مثال فاکتورهای رونویسی شامل موارد دوم است. آنها مسئول تنظیم cAMP واسطه رونویسی ژن هستند ، و یک پاسخ سلولی با تأخیر به سیگنال ارائه می دهند. علاوه بر فعال شدن پروتئین کینازها ، اردوگاه مستقیماً در کانال های غشای کاتیونی عمل می کند ، که نقش مهمی دارند ، به ویژه ، در عملکرد نورون ها. بنابراین ، سیگنال ناشی از cAMP باعث ایجاد زنجیره ای از تغییرات بیوشیمیایی در سلول هدف می شود.
کلسیم یکی دیگر از واسطه دوم که به خوبی مورد مطالعه قرار گرفته است Ca2 داخل سلولی است. یون های Ca 2+ به روش های مختلف وارد سیتوپلاسم می شوند: در امتداد کانال های غشایی (وابسته به پروتئین های G ، وابسته به ولتاژ ، تنظیم شده توسط K + یا Ca-Ca 2+) ، و همچنین از طریق کانال هایی که در مناطق ویژه ای از شبکه آندوپلاسمی قرار دارند و تحت عمل قرار می گیرند IF 3 و در عضله اسکلتی در نتیجه دپلاریزاسیون غشاء. دفع کلسیم از پلاسما سیتوزولی به دو روش انجام می شود: توسط رتیکولوم آندوپلاسمی جذب می شود و یا از سلول دفع می شود. Ca 2+ سیگنال ها را به تعداد بسیار بیشتری پروتئین نسبت به cAMP منتقل می کند - آنزیم هایی که در متابولیسم سلولی ، پروتئین کینازها ، پروتئین های اتصال دهنده کلسیم قرار دارند. دومی با سایر تأثیرگذارهای نهایی و متوسط \u200b\u200bارتباط برقرار می کنند.
تنظیم گیرنده
گیرنده ها نه تنها عملکردهای فیزیولوژیکی و بیوشیمیایی را کنترل می کنند ، بلکه به عنوان هدف تنظیم عمل می کنند. این تنظیم در سطح سنتز و تجزیه ماکرومولکولهای آنها از طریق تشکیل پیوندهای کووالانسی با سایر مولکولها ، تعامل با پروتئین های نظارتی و حرکت گیرنده انجام می شود. پروتئین های تبدیل شده و پروتئین های مؤثر نیز در معرض مقررات قرار دارند. سیگنالهای نظارتی می توانند از مسیرهای انتقال داخل سلولی فعال شده و با تحریک گیرنده خود (از طریق مکانیسم بازخورد) و همچنین از گیرنده های دیگر (مستقیم یا غیرمستقیم) فعال شوند.
تحریک طولانی مدت گیرنده های دارویی معمولاً منجر به کاهش واکنش نسبت به آن می شود - در همان غلظت ، داروی اثر کمتری را نشان می دهد. این پدیده ، به نام حساس کننده ، نسوز ، اعتیاد آور ، نقش مهمی در عمل بالینی دارد: به عنوان مثال ، با استفاده طولانی مدت آگونیست های β- آدرنرژیک برای درمان بیماران مبتلا به AD از شدت واکنش به این داروها کاسته می شود.
حساسیت زدایی هومولوژیک فقط مربوط به گیرنده های تحریک شده است و مخصوص لیگاند است. با حساسيت زدايي هترولوگ ، شدت واكنش به ساير ليگان ها ، كه گيرنده هاي آنها از طريق همين مسير انتقال سيگنال درون سلولي عمل مي كنند ، كاهش مي يابد. در حالت اول ، بازخورد منفی با تأثیرگذاری بر روی گیرنده (فسفوریلاسیون ، پروتئولیز ، کاهش سنتز) ارائه می شود ، در مورد دوم ، علاوه بر گیرنده ، می تواند بر پروتئین های دیگری که در انتقال سیگنال درون سلولی نقش دارند نیز تأثیر بگذارد.
در مقابل ، اگر گیرنده ها برای مدت طولانی تحریک نشوند ، حساسیت آنها به آگونیست ها افزایش می یابد (به عنوان مثال ، با درمان طولانی مدت با مهار کننده مهار کننده β- آدرنرژیک پروپرونولول ، حساسیت گیرنده های β-آدرنرژیک نسبت به β آدرنرژیک ها افزایش می یابد).
اختلالات ناشی از اختلال در عملکرد گیرنده
علاوه بر تفاوت های فردی در حساسیت به دارو ، بیماری هایی وجود دارد که ناشی از اختلال در عملکرد برخی از اجزای ساز و کار انتقال سیگنال درون سلولی از گیرنده به مؤثر است. با از دست دادن عملکرد گیرنده های بسیار تخصصی ، ممکن است تظاهرات فنوتیپی این بیماری محدود باشد (برای مثال ، با زنانگیزی شدن بیضه همراه با عدم وجود ژنتیکی یا نقص ساختاری گیرنده های آندروژن). اگر مکانیسم جهانی تری در انتقال سیگنال سلولی نقض شود ، علائم این بیماری متنوع تر است ، به عنوان مثال ، با میاستنی گراویس و برخی از اشکال دیابت مقاوم به انسولین ناشی از اختلالات خود ایمنی گیرنده های N-کولینرژیک و گیرنده های انسولین. نقص در هر مؤلفه ای که در انتقال سیگنال از بسیاری از گیرنده ها نقش دارد ، منجر به اختلالات غدد درون ریز می شود. به عنوان مثال شکل هتروزیگوت کمبود پروتئین G است که آدنیلات سیکلاز را در کلیه سلولها فعال می کند (اشپیگل و وینشتین ، 1995). یک شکل کمبود این پروتئین به احتمال زیاد منجر به مرگ خواهد شد.
اختلال در ساختار یا بومی سازی گیرنده ها می تواند به عنوان یک واکنش ضعیف یا تقویت شده به دارو و همچنین اثرات نامطلوب دیگر آشکار شود.
جهش دهنده های گیرنده ژن قادر به تغییر هر دو پاسخ به استفاده یگانه از دارو و همچنین اثربخشی درمان طولانی مدت هستند. به عنوان مثال ، نقص در گیرنده های β- آدرنرژیک مسئول آرامش عضلات صاف برونش ها و تنظیم مقاومت هوایی باعث کاهش حساسیت این گیرنده ها به β آدرنرژیک ها در طول مدت طولانی درمان بیماران مبتلا به AD می شود. از آنجا که جهش های مسئول عملکرد گیرنده اختلال شناسایی شده و ژن های مربوطه کلون گذاری می شوند ، می توان روش هایی برای درمان چنین بیماری هایی ایجاد کرد.
طبقه بندی گیرنده
به طور سنتی ، گیرنده های مواد مخدر بر اساس اثرات و فعالیت نسبی آگونیست های انتخابی (محرک) و آنتاگونیست ها (مسدود کننده ها) که بر روی این گیرنده ها فعالیت می کنند ، شناسایی و طبقه بندی شده اند. به عنوان مثال ، اثرات استیل کولین ، که هنگام تعامل با گیرنده های کولینرژیک آلکالوئید موسکارین مسدود می شوند و توسط آتروپین مسدود می شوند ، اثرات موسکارینی نامیده می شوند و اثرات بازتولید شده هنگام تعامل با گیرنده های کولینرژیک نیکوتین ، اثرات نیکوتینی نامیده می شوند. گیرنده هایی که واسطه اثرات موسکارین و نیکوتین هستند به ترتیب گیرنده های کولینرژیک M و N نامیده می شوند. اگرچه چنین طبقه بندی معمولاً مکانیسم عملکرد داروها را نشان نمی دهد ، اما برای سیستماتیک کردن اثرات آنها راحت است. در واقع ، این ادعا که یک دارو گیرنده های نوع خاصی را تحریک می کند ، در عین حال طیف اثرات این دارو و موادی را که باعث تقویت یا تضعیف این اثرات می شود ، تعیین می کند. اما ، اعتبار چنین ادعاهایی با شناسایی انواع جدید و زیرگروه های گیرنده ها ، کشف مکانیسم های اضافی عملکرد داروها یا عوارض جانبی قبلاً ناشناخته می تواند تغییر کند.
زیرگروه های گیرنده
با ظهور انواع فزاینده ای از داروهای بسیار منتخب ، مشخص شده است که انواع گیرنده های شناخته شده قبلاً به زیرگروه های زیادی تقسیم می شوند. روش های کلونینگ مولکولی در مطالعه زیرگروه های جدید گیرنده کمک مهمی شده است و تهیه گیرنده های نوترکیب باعث ایجاد داروهایی شده است که بصورت انتخابی روی این گیرنده ها عمل می کنند. انواع مختلفی از گیرنده های مرتبط اما غالباً (البته نه همیشه) با آگونیست ها و آنتاگونیست های مختلف ارتباط برقرار می کنند. گیرنده هایی که هیچ آگونیست انتخابی یا آنتاگونیست مشخص نشده اند ، معمولاً متعلق به یک زیرگروه واحد نیستند ، بلکه به ایزوفرم های همان گیرنده هستند. زیر گروه های جداگانه همچنین می توانند در مکانیسم های انتقال سیگنال درون سلولی متفاوت باشند. گیرنده های کولینرژیک M 1 و M3 ، به عنوان مثال ، از طریق پروتئین G q فعال می شوند ، که فسفولیپاز C را فعال می کند ، به طور غیر مستقیم باعث آزاد شدن Ca 2+ از انبارهای داخل سلولی می شود و گیرنده های کولینرژیک M2 و M4 از طریق پروتئین G i ، که مانع از آدنیلات سیکلاز می شود. در عین حال ، تقسیم گیرنده ها به انواع و زیرگروه ها اغلب نه با مکانیسم عمل بلکه با انتخاب تصادفی تعیین می شود یا براساس ایده های مستقر است. بنابراین ، گیرنده های α 1 - ، α2 - و β- آدرنرژیک در پاسخ به داروها و انتقال سیگنال متفاوت هستند (پروتئین های G i ، G q و G را به ترتیب فعال کنید) ، اگرچه گیرنده های α و β- آدرنرژیک انواع مختلفی دارند ، و α 1 - و α2- گیرنده گیرنده - به زیرگروه های مختلف در همان نوع. ایزوفرمهای α1-adrenoreceptors α 1A ، α 1B و α 1D در خصوصیات بیوشیمیایی آنها کمی متفاوت است. همان مشخصه زیر گروه های ایزوفرماز گیرنده های β- آدرنرژیک (β1 ، β2 و β 3) است.
برای ایجاد داروهای بسیار انتخابی از تفاوت هایی بین زیرگروه های گیرنده استفاده می شود ، به عنوان مثال داروهایی که به دلیل اتصال به زیرگروه های گیرنده که در مکانیسم انتقال سیگنال داخل سلولی متفاوت هستند ، تأثیرات متفاوتی در همان بافت دارند. علاوه بر این ، داروها می توانند سلولها یا بافتهای خاصی را که بیانگر گیرندههای یک زیرگروه است ، بطور انتخابی هدف قرار دهند. هرچه انتخاب داروها بیشتر باشد (در رابطه با یک بافت خاص یا در رابطه با یک اثر خاص) ، نسبت به مزایا و اثرات نامطلوب آن مطلوب تر است.
با استفاده از روش های ژنتیکی مولکولی ، نه تنها ایزوفرم های مختلف گیرنده ها کشف شده اند ، بلکه ژن هایی که گیرنده های جدید و قبلاً ناشناخته را نیز رمزگذاری می کنند ، کشف شده اند. بسیاری از این گیرنده ها قبلاً به یک یا کلاس شناخته شده دیگر اختصاص داده شده اند و عملکرد آنها با استفاده از لیگاند های مربوطه مورد مطالعه قرار گرفته است. با این حال ، هنوز برای برخی گیرنده ها لیگاند پیدا نشده است.
کشف بسیاری از ایزوفرمهای گیرنده یکسان رمزگذاری شده توسط ژنهای مختلف (خصوصاً اگر ایزوفرمها در مکانیسم انتقال سیگنال درون سلولی تفاوت نداشته و با همان لیگاندهای درون زا تعامل داشته باشند) اجازه می دهد تا بیان گیرنده ها در سلولهای مختلف مطابق با نیاز بدن در متفاوت تنظیم شود. دوره های سنی
اقدام دارویی با واسطه غیر گیرنده
همه داروها از طریق ساختارهای ماکرومولکولی - گیرنده ها عمل نمی کنند. بعضی از داروها با مولکولهای کوچک یا یونهایی که به طور طبیعی یا در یک یا وضعیت پاتولوژیک دیگر در بدن وجود دارند تعامل دارند. بنابراین ، آنتی اسیدها اسید هیدروکلریک موجود در معده را خنثی می کنند. مسنا (دارویی که به سرعت توسط کلیه ها دفع می شود و رادیکال های آزاد را خنثی می کند) به متابولیت های فعال برخی از داروهای ضد سرطان متصل می شود و باعث کاهش شدت عوارض جانبی دستگاه ادراری می شود. تعدادی از مواد بیولوژیکی غیرفعال (به عنوان مثال مانیتول) می توانند در مقادیر کافی برای افزایش اسمولاریت مایعات بیولوژیکی معرفی شوند و بدین ترتیب توزیع مایعات خارج سلولی و درون سلولی تغییر کنند. با کمک این مواد می توان دیورز را افزایش داد ، سلولهای بی سی سی را افزایش داد ، ورم مغزی را از بین برد. علاوه بر این ، از آنها به عنوان ملین استفاده می شود.
برخی از داروها به دلیل شباهت ساختاری با موادی که این اجزا را تشکیل می دهند می توانند در اجزای سلول ادغام شوند و عملکرد آنها را تغییر دهند. به عنوان مثال ، آنالوگ های پورین و پیریمیدین ها در اسیدهای نوکلئیک وارد می شوند و به عنوان ماده ضد ویروسی و ضد توموری مورد استفاده قرار می گیرند.
A.P. ویکتوروف "داروسازی بالینی"
به عنوان یک قاعده ، مکانیسم عملکرد داروها مبتنی بر توانایی آنها برای شروع (تحریک) فرآیندهای پیچیده بیوشیمیایی n / یا بیوفیزیکی است که در نهایت باعث تغییر و یا بهینه سازی فعالیت عملکرد سلول هدف می شوند.
داروها می توانند عمل خود را علیه اعضای بدن و / یا سلولهای هدف انجام دهند:
تعامل شیمیایی مستقیم؛
تعامل فیزیکی و شیمیایی بر روی غشای سلولی.
اقدامات روی آنزیم های تخصصی؛
اقدامات روی ژنهای نظارتی؛
اقدامات روی گیرنده های خاص.
تعامل مستقیم شیمیایی LS. این مکانیسم عملکرد داروها بسیار نادر است و می تواند در خارج از سلول ، به عنوان مثال ، در لومن معده یا روده ها تحقق یابد. ذات آن در این واقعیت نهفته است که داروها با یک مولکول و / یا یونهایی که در بدن در حالت عادی ایجاد می شوند با ایجاد یک وضعیت آسیب شناختی ، وارد یک واکنش شیمیایی مستقیم می شوند. نمونه ای از اثر متقابل شیمیایی مستقیم ، واکنش شیمیایی خنثی کردن اسید هیدروکلریک معده هنگام مصرف داروهای ضد اسید است (نگاه کنید به ج 2 ، ص 112).
اثر متقابل فیزیکی و شیمیایی داروها بر روی غشای سلولی. یکی از کارکردهای اصلی غشای سیتوپلاسمی اجرای تبادل یونی بین سیتوپلاسم و محیط خارج سلولی است. تبادل یونی transmembrane همچنین می تواند از طریق کانال های یون یونی وابسته به ولتاژ - سدیم ، پتاسیم ، کلسیم ، کلر و غیره انجام شود. برخی از داروها با رسیدن به غشای سلولی با این کانالها ارتباط برقرار می کنند و فعالیت عملکردی آنها را تغییر می دهند. بنابراین ، به عنوان مثال ، عمل ضد آریتمی یک داروی کلاس IA ، کینیدین ، \u200b\u200bمبتنی بر توانایی آن در مسدود کردن عبور یون های Na + از طریق کانال های سدیم ترانسمیبنی است (نگاه کنید به ج 2 ، ص 35).
تأثیر داروها بر آنزیمهای تخصصی. مقدار نسبتاً کمی از داروها با تغییر فعالیت برخی از آنزیمهای سلولی تخصصی ، اثر دارویی آن را درک می کنند. داروهایی که باعث افزایش فعالیت آنزیم های سلولی می شوند ، القاکننده آنزیم نامیده می شوند. چنین عملی ، به عنوان مثال ، با قرص های خواب آور و داروی ضد تشنج فنوباربیتال ، که به طور قابل توجهی فعالیت آنزیم های کبدی میکروزومی را تقویت می کند ، وجود دارد. اهمیت بیولوژیکی این اثر فنوباربیتال و نزدیک به آن LS در زیر مورد توجه قرار خواهد گرفت.
داروهای مهار کننده فعالیت آنزیم های تخصصی به عنوان مهارکننده های آنزیمی گفته می شوند. بنابراین ، به عنوان مثال ، داروی ضد افسردگی از گروه مهار کننده های مونوآمین اکسیداز (MAOs) ، داروی پیرلیندول با مهار فعالیت آنزیم MAO در سیستم عصبی مرکزی ، اثر ضد افسردگی خود را تحقق می بخشد (نگاه کنید به تی 1 ، ص 294).
توانایی مهار فعالیت آنزیم استیل کولین استراز اساس فعالیت دارویی داروهای آنتی کولین استراز ، به عنوان مثال فیزیوستیگمین است. مشخص شده است که در شرایط فیزیولوژیکی ، استیل کولین استراز غیرفعال می شود (از بین می برد) استیل کولین ، انتقال دهنده عصبی است که باعث تحریک تحریک در سیناپس های سیستم عصبی پاراسمپاتیک می شود. Physostigmine ، با سرکوب فعالیت استیل کولین استراز ، باعث افزایش تجمع در سیناپسهای سیستم پاراسمپاتیک از استیل کولین انتقال دهنده عصبی می شود ، در نتیجه ، میزان صدای سیستم عصبی پاراسمپاتیک افزایش می یابد ، که در سطح سیستمیک بواسطه ایجاد برادی کاردی ، کاهش فشار خون و فشار خون (فشار خون افزایش می یابد) مردمک و غیره
داروها می توانند با آنزیم ها برگشت پذیر و برگشت ناپذیر داشته باشند. به عنوان مثال ، داروی انالاپریل به طور معکوس فعالیت آنزیم تبدیل کننده آنژیوتانسین را مهار می کند ، که به ویژه به کاهش فشار خون منجر می شود ، در حالی که مواد سمی ارگانوفسفر باعث برگشت ناپذیر فعالیت استیل کولین استراز می شوند.
تأثیر داروها بر روی ژنهای نظارتی. در حال حاضر ، دانشمندان در تلاشند تا داروهایی را ایجاد کنند که اثرات دارویی آنها را با تأثیر مستقیم بر فعالیت فیزیولوژیکی ژنهای نظارتی تأثیر بگذارد. به نظر می رسد این روند پس از کشف ساختار ژنوم انسانی در سال 2000 امیدوار کننده است. اعتقاد بر این است که عادی سازی انتخابی عملکرد ژنهای نظارتی تحت تأثیر داروها باعث می شود که در درمان بسیاری از بیماریها از جمله بیماریهایی که قبلاً غیرقابل تحمل بوده اند ، موفقیت حاصل شود.
تأثیر داروها بر روی گیرنده ها. قبل از اینکه به ویژگیهای تعامل داروها با گیرنده ها بپردازیم ، لازم است منظور ما را از اصطلاح گیرنده مشخص کنیم (از لاتین receio) (از لاتین receio) استفاده کنید.
از دوره فیزیولوژی مشخص شده است که اصطلاح گیرنده به معنای سازندهای کاملاً تخصصی است که قادر به درک ، تبدیل و انتقال انرژی یک سیگنال خارجی به سیستم عصبی هستند. این گیرنده ها را حسی (از لاتین. نظرسنجی - احساس ، احساس ، ادراک) می نامند.
گیرنده های حسی شامل گیرنده های اندام های شنوایی ، بینایی ، بو ، بو ، مزه ، لمس و غیره هستند. گیرنده های حسی این اندام ها متعلق به به اصطلاح بیرون گیرنده ها هستند.
اگر از زمان های قدیم وجود ارگان های حسی که به محرک های خارجی تحریک پاسخ می دهند ، وجود گیرنده های حسی در داخل بدن مورد سوال قرار گرفته است. برای اولین بار ، وجود چنین گیرنده هایی در داخل بدن توسط فیزیولوژیست روسی I.F.Pion پیشنهاد شد که در سال 1866 به دلیل تحریک آئورت در یک آزمایش خرگوش افت فشار خون را نشان داد. این کشف باعث جستجو و مطالعه گیرنده های مستقر در بدن شد و خود این گیرنده ها به عنوان گیرنده گیرنده نامیده شدند.
با آغاز قرن بیستم تعداد کافی از گیرنده های حسی آشکار شد و نقش مهم آنها در تنظیم عملکردهای فیزیولوژیکی بدن ثابت شد.
در سال 1905 ، J. Langley ثابت کرد که هنگامی که یک دارو بر روی غشای سلولی اعمال می شود ، اگر فقط در یک منطقه خاص از آن استفاده شود ، اثر دارویی ایجاد می شود. علاوه بر این ، این سایت تنها بخش کوچکی از مساحت کل سطح سلول را تشکیل می دهد. این مشاهده به J. Langley اجازه داد که نتیجه گیری کند که سایت های مختلف گیرنده در تعامل با داروها بر روی غشای سلولی وجود دارند.
اما اولویت در ایجاد تئوری گیرنده عمل به داروها متعلق به فیزیولوژیست آلمانی P. P. Erlich است که در سال 1906 برای اولین بار اصطلاح "گیرنده" را معرفی کرد و فرمولاسیون فرمولاسیون را عنوان کرد "در صورت ثابت بودن بر روی غشای سلولی ، دارو کار نمی کند." طبق تئوری P. Ehlich ، یک مولکول دارویی دارای دو گروه عملکردی فعال است که یکی از آنها تثبیت آن بر روی سطح سلول در ناحیه گیرنده دارو را تضمین می کند ، و گروه دوم عملکردی با گیرنده ارتباط برقرار می کنند و زنجیره پیچیده ای از واکنشهای بیوشیمیایی را ایجاد می کنند که فعالیت فیزیولوژیکی آن (سلول) را تغییر می دهد. .
بنابراین ، در اوایل قرن بیستم. مشخص شد که حداقل دو کلاس گیرنده گیرنده وجود دارد: گیرنده های حسی که اطلاعات مربوط به وضعیت اندام های داخلی و بافت های بدن را به سیستم عصبی مرکزی منتقل می کنند. گیرنده های برچسب زدن با داروهایی که فعالیت عملکردی سلولهای هدف را تغییر می دهند تعامل دارند.
بلافاصله لازم به ذکر است که در آینده ، در متن کتاب درسی ، برای جلوگیری از سردرگمی در اصطلاحات ، گیرنده های مواد مخدر و مواد بیولوژیکی فعال ، یعنی. دارای برچسب ، یا گیرنده های گشایی با استفاده از اصطلاح "گیرنده" مشخص می شود ، در حالی که گیرنده های حسگر با اصطلاح مشخص کننده فعالیت های عملکردی خود ، به عنوان مثال "گیرنده های گیرنده" ، "گیرنده های درد" و غیره مشخص می شوند.
کشف P. Erlich در غشای سلولی گیرنده های مواد مخدر به عنوان نقطه شروع برای توسعه علم داروسازی ، به ویژه فارماکودینامیک ، که یکی از اصلی ترین وظایف آن مطالعه مکانیسم های گیرنده عملکرد داروها است.
در حال حاضر ، ساختار تعداد زیادی گیرنده سلولی ، ویژگیهای متقابل برخی از ترکیبات فعال بیولوژیکی با آنها آشکار شده است ، که این امر باعث می شد از یک طرف درک مکانیسم عملکرد داروهای شناخته شده و از طرف دیگر زمینه ساز ایجاد داروهای جدید بسیار مؤثر باشد.
به طور طبیعی ، دشوار است تصور کنید که در جریان تکامل ، گیرنده های داروهای مختلف مصنوعی (شیمیایی به دست آمده) در بدن انسان شکل گرفته است ، به خصوص که اکثریت قریب به اتفاق داروهای ارائه شده در بازار دارویی مدرن در 50 سال گذشته یا کمتر سنتز شده اند. ثابت شده است که دستگاه گیرنده سلول یک سازه کاربردی-ساختاری بسیار باستانی است. بنابراین ، گیرنده های a- و β (گیرنده ها ، اثر متقابل نوراپی نفرین و آدرنالین بر عملکرد عملکرد سلول تأثیر می گذارد) نه تنها در سلول های حیوانی بلکه در غشاهای سلولی سلول های گیاهی نیز مشاهده می شود ، به عنوان مثال در سلول های گیاه نیتلا ، جایی که a- و β- آدرنرژیک حرکت پروتوپلاسم (محتوای سلول) را تنظیم می کند.
سپس گیرنده های داروهای کشف شده توسط P. Ehrich چه هستند ، و چرا آنها با آنها ارتباط برقرار می کنند؟
در حال حاضر ، شکی نیست که گیرنده های به اصطلاح دارویی در واقع گیرنده هایی برای مواد فعال بیولوژیکی درون زا (تولید شده در بدن) هستند که در تنظیم فعالیت عملکردی اندام های داخلی و بافت های بدن نقش دارند. چنین ترکیبات فعال بیولوژیکی شامل موادی است که در هنگام انتقال سیگنال عصبی از انتهای عصب رها می شود ، همچنین هورمون ها ، ویتامین ها ، اسیدهای آمینه و غیره. برای هر ماده بیولوژیکی فعال درون زا ، گیرنده های خاصی برای آن وجود دارد. بنابراین ، به عنوان مثال ، ماده بیولوژیکی فعال تولید شده در بدن ، آدرنالین ، می تواند بطور جدی خاص a- و β- آدرنرفتورها را فعال کند ، و گلوکوکورتیکواستروئیدها - هورمونهای قشر آدرنال - تنها با گیرنده های گلوکوکورتیکوئیدوئید به طور خاص برای آنها ارتباط برقرار کنند.
داروهای مصنوعی که با تعامل با دستگاه گیرنده سلول ، اثرات خود را متوجه می شوند ، در ساختار شیمیایی خود ، کم و بیش به ترکیبات بیولوژیکی فعال درون زا که با گیرنده های مشابه تعامل دارند ، شباهت دارند. به عنوان مثال ، داروهای فنیل افرین (وازو کانستریکتور مصنوعی) (باعث انقباض عروق) می شوند و فنیل افرین در ساختار شیمیایی خود با ماده فعال درون زا فعال بیولوژیکی نوراپی نفرین نزدیک است ، بنابراین ، مانند نوراپی نفرین ، توانایی تحریک آدرنرژیک ها را دارد.
بعضی اوقات ، به دلیل ویژگی های ساختار شیمیایی آنها ، داروها می توانند نه با خود گیرنده بلکه با قسمت مجاور غشای سلولی ارتباط برقرار کنند. از آنجا که در این حالت ، دارو با گیرنده خود تعامل ندارد ، اما با قسمت مجاور غشای سلولی ، آنها از یک اثر هیجان انگیز یا مسدود کننده بر روی گیرنده صحبت نمی کنند ، بلکه از یک ماده آلوستریک (از alios یونانی - اثر دیگر ، متفاوت) یا اثر است. در نتیجه ممکن است تغییری در ساختار غشای مجاور گیرنده و اجزای فردی گیرنده ایجاد شود که ممکن است منجر به تغییر حساسیت گیرنده به یک ماده بیولوژیکی فعال مخصوص آن شود. در مواردی که حساسیت گیرنده به یک ماده فعال بیولوژیکی افزایش می یابد ، آنها از حساسیت (از sensus لاتین - حس) یا حساسیت (از sensinilis لاتین - حساسیت) گیرنده صحبت می کنند و در مواردی که حساسیت گیرنده کاهش می یابد ، از حساسیت زدایی صحبت می کنند. گیرنده
ویژگی خاصیت آلوستریک در این واقعیت نهفته است که داروهایی که دارای این نوع مکانیسم عمل هستند مستقیماً در انتقال یک ضربه عصبی تأثیر نمی گذارند بلکه آن را در جهت مورد نظر اصلاح می کنند. به عنوان مثال ، مکانیسم عمل ضد اضطراب (داروهای ضد اضطراب ؛ مترادف: آرامبخشها) که در ساختار شیمیایی آنها مشتقات بنزودیازپین هستند ، مبتنی بر پدیده تحریک آلوستریک گیرنده های بنزودیازپین پس سیناپسی است. تحریک دوم ، به نوبه خود ، باعث تقویت فعال شدن گیرنده های پس سیناپسی مهار کننده اسید گاما آمینوبوتیریک (گیرنده های گابا GABA) می شود که با از بین بردن علائم بیماری های عصبی مانند اضطراب ، اضطراب ، ترس و غیره از نظر بالینی تجویز می شود.
گیرنده ها ، در تعامل با آنها ماده یا دارویی بیولوژیکی فعال به هر طریق حالت عملکردی سلول هدف را تغییر می دهند ، خاص نامیده می شوند.
علاوه بر گیرنده های خاص ، به اصطلاح گیرنده های مخصوص دارو جدا شده اند. در ادبیات تخصصی پزشکی ، به این گیرنده ها "محل از بین رفتن" داروها نیز گفته می شود. با تماس با چنین گیرنده ها ، داروها هیچ اثر بیولوژیکی ندارند ، اما خود آنها بیولوژیکی غیرفعال می شوند. نمونه ای از این نوع گیرنده می تواند به عنوان گیرنده هایی که بر روی پروتئین های پلاسما قرار دارند ، بخصوص پروتئین های محلول در آب - آلبومین - خدمت کند. اهمیت این پدیده در زیر به تفصیل مورد بحث قرار خواهد گرفت (به تی 1 ، ص 72 مراجعه کنید).
ساختار گیرنده ها کاملاً پیچیده است ، اما بیشتر آنها پروتئین ماکرومولکول ها یا گلیکوپروتئین ها هستند که ممکن است شامل یون ها ، لیپیدها ، اسیدهای نوکلئیک و غیره نیز باشند. گیرنده یعنی. ماکرومولکول پروتئین تشکیل دهنده آن با استفاده از یک ترکیب خاص و خاص برای هر گیرنده ، ترتیب مکانی گروههای شیمیایی آن است. ماکرومولکول پروتئین تشکیل دهنده گیرنده می تواند در لایه لیپید غشای سیتوپلاسمی یا در داخل سلول بصورت یکپارچه (غوطه ور) شود. کارکرد اصلی گیرنده سلولی این است که یک سیگنال شیمیایی که از طریق یک ماده فعال بیولوژیکی فعال درون زا و یا داروها به آن منتقل می شود را "بشناسد" و آن را به پاسخ بیوشیمیایی و / یا بیوفیزیکی مربوطه تبدیل کند.
پیش از این اعتقاد بر این بود که داروها یا مواد بیولوژیکی فعال درون زا با گیرنده های نوع "کلید و قفل" ارتباط برقرار می کنند ، یعنی. گیرنده دارای ساختاری است که به دارو اجازه می دهد تا گیرنده "شما" را پیدا کند ، به آن متصل شود و مانند گذشته "روشن" شود و "خاموش" شود. با این حال ، با پیشرفت علم پزشکی ، آشکار شده است که نفس کاملاً اینگونه نیست. در حال حاضر ، فرآیندهای مولکولی تبدیل سیگنالهای خارج سلولی به عملکرد سلولی تنظیم کننده داخل سلولی ، قبلاً کاملاً مورد مطالعه قرار گرفته است. مکانیسم هایی که منجر به اثر متقابل مواد درون زا و بیولوژیکی فعال مواد مخدر با گیرنده ها می شوند.
هنگام تعامل با گیرنده یک ماده بیولوژیکی فعال درون زا و / یا یک L فعال C مانند آن ، یک ترکیب ایجاد می شود - یک تغییر مکانی به شکل یک ماکرو مولکول پروتئین ، که ماشه ای برای فرآیندهای درون سلولی مختلف است که پاسخ سلول هدف به یک واسطه و / یا دارو را تعیین می کند. به عنوان مثال ، فعال شدن گیرنده های آدرنرژیک عضله صاف برونش تحت تأثیر فنوترول β2-آدنوستیمولاتور منجر به افزایش فعالیت آنزیم آدنیلات سیکلاز ، که به تجمع مونوفسفات آدنوزین حلقوی (cAMP) در سلول و در نتیجه آرام سازی سلول منجر می شود.
به طور کلی از نظر بیولوژیکی ، گیرنده های سلولی می توانند به عنوان "ارگان های حسی" کاملاً تخصصی در نظر گرفته شوند که از طریق آنها "اطلاعات" را که مثلاً از سیستم عصبی مرکزی و / یا سیستم غدد درون ریز ناشی می شود ، درک می کنند. با وجود نقش مهم دستگاه گیرنده ، گیرنده ها تنها بخش ناچیزی از غشای سلولی را اشغال می کنند. به عنوان مثال ، دستگاه گیرنده M- کولینرژیک یک سلول بیش از 1/6000 مساحت آن را اشغال نمی کند.
بررسی خصوصیات تعامل داروها با گیرنده ، از یک سو به ما این امکان را می دهد تا اساس مکانیسم مولکولی عمل آن را بفهمیم و از طرف دیگر ، اطلاعاتی را در مورد اینکه چه عواملی باید در ساختار داروها ایجاد شود را فراهم می کند تا توانایی آن در تعامل با این گیرنده را تقویت کند ، یعنی. . سنتز هدفمند داروهای جدید بسیار مؤثر را امکان پذیر می کند.
تحت شرایط فیزیولوژیکی ، گیرنده های مختلف سلولی به طور مستقل عمل نمی کنند ، اما در تعامل مداوم با یکدیگر هستند و از این طریق فعالیت خاص سلول را تنظیم می کنند. به عنوان مثال ، فعال شدن گیرنده های β- آدرنرژیک قلبی توسط نوراپی نفرین درون زای باعث بخصوص افزایش تعداد انقباضات قلبی و فعال شدن گیرنده های M- کولینرژیک سلول های قلبی توسط استیل کولین درون زا ، برعکس باعث کاهش تعداد انقباضات قلب می شود.
با کشف گیرنده های پیش و پس از سیناپسی ، سهم بسزایی در درک مکانیسم های گیرنده عمل مواد مخدر داشت. سیناپس (از synapsis یونانی - اتصال ، ارتباطات) یک منطقه تماس تخصصی بین سلولهای عصبی یا سایر ساختارهای تحریک آمیز بدن است که انتقال اطلاعات ورودی و حفظ اهمیت اطلاعاتی آن را تضمین می کند. مطالعه ساختار و نقش عملکردی سیناپسها در اواخر قرن XIX آغاز شد. پس از آن ، بافت شناس اسپانیایی S. Ramon n Cajal (S. Ramon در Cajal) حضور سیستم انتقال تخصصی در سیستم عصبی مرکزی را پیشنهاد کرد. سیناپس ها نام خود را در سال 1897 بدست آوردند ، زمانی كه فیزیولوژیست انگلیسی C. Sherrington این اصطلاح را پیشنهاد كرد كه به ناحیه تماس بین سلولهای عصبی مراجعه كند.
در حال حاضر ، سه نوع سیناپس وجود دارد:
1) سیناپسهای "الکتریکی" که در آن اطلاعات با انتقال سیگنال الکتریکی از غشای پیش سیناپسی منتقل می شود. به این نوع سیناپس efaps گفته می شود (از یونانی. Ephapsis - تماس تنگ).
2) سیناپسهای "شیمیایی" که در آن اطلاعات از طریق مواد فعال بیولوژیکی ویژه منتقل می شود - انتقال دهنده های عصبی (از یونانی. نورون - عصب و لاتین. واسطه - واسطه؛ مترادف: واسطه).
3) سیناپسهای "مختلط" که در آن اطلاعات به صورت شیمیایی و برقی منتقل می شوند.
اثرات دارویی اکثریت قریب به اتفاق داروهایی که بر عملکرد سیناپسها تأثیر می گذارند ، با تأثیر آنها بر گوات یا مرحله دیگری از انتقال سیگنال در سیناپسهای شیمیایی ، یعنی متوجه می شوند. در خلاصه نوع دوم.
به عنوان یک قاعده ، سیناپسهای شیمیایی توسط انتقال دهنده های عصبی طبقه بندی می شوند که تکانه های عصبی را در آنها منتقل می کنند ، به شرح زیر است:
سیناپسهایی که استیل کولین به عنوان واسطه عمل می کنند ، کولینرژیک نامیده می شوند.
سیناپسهایی که آدرنالین و نوراپی نفرین به عنوان واسطه عمل می کنند ، آدرنرژیک نامیده می شوند.
سیناپسهایی که در آنها ATP و آدنوزین به عنوان واسطه عمل می کنند ، پورینرژیک نامیده می شوند.
سیناپسهایی که در آنها گاما آمینوبوتیریک اسید به عنوان واسطه عمل می کند ، GABA-ergic و غیره نامیده می شوند.
ساختار سیناپس در حال حاضر به خوبی درک شده است. سیناپس شامل یک فرآیند پیش سیناپسی سلول عصبی (انتهای آکسون) و دستگاهی برای دریافت سیگنال است که بر روی غشای یک سلول مؤثر ("اجرایی") قرار دارد.
آکسون نورون efferent ، با نزدیک شدن به سلول مؤثر ، غلاف میلین را از دست می دهد ، به اصطلاح ضخیم شدن پیش سیناپسی را منبسط می کند و تشکیل می دهد (شکل 1.5). سطح عصبی منتهی به غشای سلولی سلول مؤثر ، غشای پیش سیناپسی نامیده می شود. محل سلول مؤثر در مقابل غشای پیش سیناپسی غشای پست سیناپسی نامیده می شود (شکل 1.5 را ببینید). بسته به ویژگی های ساختاری سیناپس ، ممبران پیش سیناپسی ممکن است دارای چین های کمابیش یا کمتری باشد و در نتیجه منطقه بزرگتر یا کوچک تری داشته باشد. در سیناپسهای شیمیایی ، غشای پیش سیناپسی به طور مستقیم با غشای پس سیناپسی تماس نمی گیرد ، بلکه با فاصله کمی به نام شکاف سیناپسی از آن جدا می شود (شکل 1.5 را ببینید).
ضخیم شدن پیش سیناپسی ، یعنی قسمت انتهایی آکسون شامل تعداد بیشتری میتوکندری ، اندامکهای درون سلول درگیر در سنتز و تجمع انرژی است که بیشتر از بدن نورون است و این نشانگر شدت فرآیندهای انرژی است که در این بخش از سلول عصبی رخ می دهد. علاوه بر میتوکندری ، ضخیم شدن پیش سیناپسی حاوی تعداد زیادی از وزیکولهای کوچک - وزیکولها است. به طور متوسط \u200b\u200b، حدود 20،000 وزیکول در یک ضخیم شدن پیش سیناپسی موجود است. دومی در ضخیم شدن پیش سیناپسی به طور ناهموار بومی سازی می شوند ، به عنوان یک قاعده ، بیشتر آنها در نزدیکی غشای پیش سیناپسی قرار دارند. انتقال دهنده عصبی در بدن و آکسون نورون ساخته می شود و در وزیکول ها تجمع می یابد. هر وزیکول حاوی چندین هزار مولکول انتقال دهنده عصبی (از I 000 تا 50 000) است. هنگامی که یک ضربه عصبی رخ می دهد ، وزیکول با غشای پیش سیناپسی فیوز می شود و انتقال دهنده عصبی در شکاف سیناپسی ترشح می شود (شکل 1.5 را ببینید).
شکل 1.5 نمودار شماتیک ساختار سیناپس "شیمیایی":
a یک تصویر شماتیک است. ب - میکروگراف الکترونیکی؛ 1- پایان دادن به عصب پیش سیناپسی؛ 2 - غشای پارس سیناپسی؛ 3 - غشای پس سیناپسی؛ 4 - شکاف سیناپسی؛ ب - وزیکول؛ NM - انتقال دهنده عصبی؛ P - گیرنده postynaptic: OZ - ضبط "معکوس" از انتقال دهنده عصبی. SF یک آنزیم تخصصی است که انتقال دهنده عصبی اضافی در شکاف سیناپسی را از بین می برد
سازندهای گیرنده فعال عملکردی بر روی غشای پست سیناپسی قرار دارند ، که قادر به تعامل با انتقال دهنده عصبی هستند که در طی عبور یک ضربه عصبی از غشای پیش سیناپسی رها شده است. گیرنده های واقع در غشای پست سیناپسی در ادبیات پزشکی تخصصی به گیرنده های سیناپسی یا پس سیناپسی گفته می شوند. با استفاده از گیرنده های پس سیناپسی به معنای ماکرومولکول های پروتئین در داخل غشای بعد از سیناپسی با ساختار و عملکرد از پیش تعیین شده ژنتیکی قرار گرفته است ، که قادر به تعامل معکوس با فرستنده های عصبی و / یا داروها به دلیل گروه های عملکردی مرکز فعال (قسمت "تشخیص" بخش ماکرومولکول) هستند.
انتقال سیگنال عصبی در سیناپس به شرح زیر است: تحت تأثیر یک محرک عصبی ، وزیکول ها به غشای پیش سیناپسی منتقل می شوند و انتقال دهنده عصبی توسط اگزوسیتوز به شکاف سیناپسی ترشح می شود (شکل 1.5 را ببینید). انتقال دهنده عصبی که به شکاف سیناپسی آزاد می شود به غشای پست سیناپسی می رسد ، جایی که با تعامل با گیرنده پس سیناپسی ، زنجیره ای از واکنشهای بیوشیمیایی و / یا بیوفیزیکی را تحریک می کند ، که نتیجه آن پاسخ فیزیولوژیکی سلول هدف است. با این حال ، تمام میزان انتقال دهنده عصبی منتشر نشده به گیرنده های پس سیناپسی می رسد و با آنها در تعامل است. بخشی از انتقال دهنده عصبی توسط غشای پیش سیناپسی اسیر می شود و به محل های ذخیره سازی باز می گردد. به این پدیده پدیده بازگشت مجدد نوروترانسمیتر گفته می شود.
مقدار باقیمانده گیرنده های انتقال دهنده عصبی غیر متقابل توسط آنزیم های تخصصی در شکاف سیناپسی نابود می شود. به این پدیده تخریب انتقال دهنده های عصبی گفته می شود. به عنوان مثال ، آنزیم استیل کولین استراز کاتالیز می کند (تسریع می کند) روند تخریب (تخریب) در شکاف سیناپسی از استیل کولین انتقال دهنده عصبی.
برخلاف انتقال دهنده عصبی ، محصولات متابولیکی آن دارای فعالیت انتقال دهنده عصبی هستند. کل فرآیند تعامل انتقال دهنده عصبی با گیرنده ها و از بین بردن مقدار اضافی آن توسط آنزیم خاص بسیار کوتاه است و از 2 ms (1 ms \u003d 0.001 ثانیه) تجاوز نمی کند.
چنین مدت كوتاهی از این فرآیند ، از یك سو با انتشار بسیار سریع انتقال دهنده عصبی از گیرنده و از طرف دیگر با میزان بالای غیرفعال كردن آنزیمی از انتقال دهنده عصبی در كلاه سیناپسی توضیح داده شده است.
فعالیت اساسی عملکرد سیناپس به صورت زیر قابل تغییر است:
برای تسریع ، کاهش یا مسدود کردن سنتز ، انباشت و / یا کاتابولیسم (تخریب) انتقال دهنده عصبی در انتهای پیش سیناپسی. در نتیجه این ، محتوای انتقال دهنده عصبی و در نتیجه این امر ، شدت فعالیت فیزیولوژیکی آن به نوعی تغییر خواهد کرد.
به عنوان مثال ، رزرپین سمپاتولیتیک تا هنگام تخلیه کامل آنها از تجمع کاتکول آمین ها در وزیکول سیناپسی جلوگیری می کند. در نتیجه ، مقدار نوراپی نفرین عصبی منتقل شده به شکاف سیناپسی به شدت کاهش می یابد. در سطح سیستم ، این اثر در قالب کاهش فشار خون تحقق می یابد. برخی از داروها به طور مستقیم بر محتوای انتقال دهنده های عصبی در انتهای پیش سیناپسی تأثیر نمی گذارند ، اما فعالیت آنزیم هایی را که باعث از بین رفتن آنها می شوند ، مهار می کنند. بنابراین ، تعدادی از داروهای ضد افسردگی عمل می کنند. به عنوان مثال ، داروی ضد افسردگی پیرلیندول فعالیت آنزیم مونوآمین اکسیداز در خاتمه پیش سیناپسی را مهار (سرکوب می کند) و در نتیجه باعث افزایش غلظت انتقال دهنده های عصبی مانند نوراپی نفرین ، دوپامین و سروتونین در آن می شود. از نظر بالینی ، این اثر پیرلیندول با کاهش احساس اضطراب و ترس ، بهبود خلق و خو ، افزایش فعالیت بدنی و غیره آشکار می شود.
توانایی انتقال دهنده عصبی در نفوذ به غشای پیش سیناپسی را تغییر داده (تسهیل ، پیچیده تر) و بنابراین ، با هر پالس ، میزان انتقال دهنده عصبی که در شکاف سیناپسی آزاد می شود ، افزایش یا کاهش می یابد.
به عنوان مثال ، آمفتامین روانپزشکی باعث آزادسازی کاتکول آمین ها در سیناپس های آدرنرژیک سیستم عصبی مرکزی می شود و از این طریق محتوای آنها در شکاف سیناپسی افزایش می یابد. از نظر بالینی ، این اثر دارو با خلق و خوی بهبود یافته ، احساس افزایش قدرت ، افزایش کارایی نشان داده می شود. سم tetanus باعث آزاد شدن انتقال دهنده های عصبی مهاری (GABA ، گلیسین) در سیستم عصبی مرکزی می شود و از این طریق به شدت محتوای آنها را در شکاف سیناپسی کاهش می دهد ، که از نظر بالینی با ایجاد تشنج آشکار می شود.
بازگرداندن انتقال دهنده های عصبی توسط غشای پیش سیناپسی را مسدود یا تحریک کرده و بنابراین ، غلظت انتقال دهنده های عصبی در شکاف سیناپسی را افزایش یا کاهش می دهد.
به عنوان مثال ، ایمیپرامین سه داروی ضد افسردگی مجدد بازگرداندن نوروترینفرین نوروترانسمیترین توسط غشای پیش سیناپسی را مسدود می کند و از این طریق به شدت غلظت آن را در شکاف سیناپسی افزایش می دهد. از نظر بالینی ، این اثر ایمی پرامین با بهبود خلق و خو ، افزایش فعالیت های ذهنی و جسمی آشکار می شود.
فعالیت آنزیم هایی که انتقال دهنده عصبی در شکاف سیناپسی را از بین می برد تحریک یا مسدود می کند.
به عنوان مثال ، داروی آنتی کولین استراز فیزوستیگمین فعالیت آنزیم استیل کولین استراز را کاهش می دهد ، که استیل کولین عصبی انتقال دهنده عصبی در شکاف سیناپسی را از بین می برد و از این طریق به افزایش غلظت آن کمک می کند ، که می تواند از نظر بالینی خصوصاً با کاهش فشار داخل چشم و انقباض شفیره از بین رود.
گیرنده های پس سیناپسی را تحریک یا مسدود کنید ، یعنی اثر انتقال دهنده های عصبی را تقلید یا مسدود کنید.
به عنوان مثال ، مسکن های مخدر که باعث تحریک گیرنده های مواد افیونی بعد از سیناپسی می شوند و از این طریق تأثیر انتقال دهنده های عصبی - انکفالین ها را تقلید می کنند. استریچین با مسدود کردن گیرنده های گلیسین انتقال دهنده عصبی مهاری ، مانع از تحقق اثر مهاری آن می شود ؛ در نتیجه ، استریکنین در دوزهای زیاد باعث تشنج می شود.

شکل 1.6 بازنمایی شماتیک از محلی سازی گیرنده های قبل و بعد از سیناپسی که توسط سیناپس آدرنرژیک نشان داده شده است (توضیح در
NM - انتقال دهنده عصبی؛ M2 (-) - heteroreceptor پیش سیناپسی "مهاری" کولینرژیک. β 1 (+) - "فعال سازی" گیرنده تحریک کننده پیش سیناپسی آدرنرژیک. β - گیرنده postynaptic آدرنرژیک
علاوه بر گیرنده های واقع در غشای پست سیناپسی ، یعنی. گیرنده های پس سیناپسی ، گیرنده های واقع در غشای پیش سیناپسی ، یعنی. گیرنده های پیش سیناتیک (شکل 1.6). با وجود این واقعیت که گیرنده های پیش و بعد از سین سینتیک می توانند توسط یک انتقال دهنده عصبی یکسان هیجان زده شوند ، نقش عملکردی این سازندهای گیرنده در سیناپس ها متفاوت است. اگر گیرنده های پس سیناپسی پیوند نهایی انتقال یک ضربه عصبی به اندام مؤثر باشد ، یعنی هدایت یک طرفه یک تکانه عصبی از مرکز تا حاشیه را فراهم می کند ، سپس گیرنده های پیش سیناپسی در
تنظیم فعالیت انتقال دهنده عصبی سیناپس ، به عنوان مثال تا حدی فرآیندهای رهاسازی و / یا سنتز انتقال دهنده عصبی در آن را تحت تأثیر قرار می دهد. باید تأکید کرد که گیرنده های پیش سیناپسی به طور مستقیم در هدایت تکانه عصبی از یک نورون به یک اندام مؤثر شرکت نمی کنند.
گیرنده های پیش سیناپسی به دو گروه بزرگ تقسیم می شوند: گیرنده های خودکار و هتروئونولودولار (در شکل 1.6 مراجعه کنید).
گیرنده های پیش سیناپسی شامل گیرنده هایی هستند که توسط انتقال دهنده عصبی خود برای این سیناپس هیجان زده می شوند.
به عنوان مثال ، در سیناپس هایی که در ناحیه تماس بین اعصاب جسمی و عضله منقبض قرار دارند ، هنگامی که انتقال دهنده عصبی حاوی مقدار زیادی استیل کولین در شکاف سیناپسی است ، در تعامل با autorecepters پیش سیناپسی ، از انتشار بخش جدیدی از انتقال دهنده عصبی جلوگیری می کند. تحریک autoreceptors پیش سیناپسی تنظیم آزادی استیل کولین از پایانه های پیش سیناپسی را تنظیم می کند.
با این حال ، بر روی غشای پیش سیناپسی ، علاوه بر autoreceptors ، یعنی. گیرنده هایی که به انتقال دهنده عصبی حساس هستند و تحریک را در یک سیناپس معین منتقل می کنند ، می توانند گیرنده هایی قرار گیرند که به یک انتقال دهنده عصبی حساس نباشند که تحریک را در یک سیناپس معین منتقل می کند ، اما با نوع دیگری از انتقال دهنده عصبی ارتباط دارند.
به عنوان مثال ، روی غشای پیش سیناپسی سیناپسی ، که در آن استیل کولین انتقال دهنده عصبی است ، می توان گیرنده های پیش سیناپسی حساس به نوراپی نفرین عصبی انتقال دهنده عصبی قرار داد. این نوع گیرنده پیش سیناپسی را گیرنده هترونورمود کننده کننده می نامند.
بنابراین ، سیناپس یک شکل گیری پیچیده آناتومیکی و عملکردی است که انتقال نیروی عصبی از یک نورون به یک نورون یا از یک نورون به یک سلول مؤثر را تضمین می کند.
ترتیب فعالیت عملکرد سیناپس (مراحل انتقال سیناپسی) به شرح زیر است:
سنتز و تجمع یک انتقال دهنده عصبی در وزیکولهای محلی شده در ضخامتهای پیش سیناپسی (سنتز یک انتقال دهنده عصبی نه تنها در ضخیم شدنهای پیش سیناپسی بلکه در سلولهای عصبی و آکسونها اتفاق می افتد).
تركیبكننده انتقال دهنده عصبی به شکاف سیناپسی در زمان عبور یک ضربه عصبی.
تعامل انتقال دهنده عصبی با گیرنده های پس سیناپسی ، که مستلزم فعال شدن گیرنده ها و تغییر در عملکرد عملکردی سلول مؤثر است.
غیرفعال کردن انتقال دهنده عصبی (آنزیمی) و / یا بازگرداندن مجدد آن توسط غشای پیش سیناپسی ، یعنی. ترمیم توانایی سیناپس برای انتقال مجدد عصب به سلول مؤثر.
سیناپسها دارای خصوصیات اساسی زیر هستند:
انجام یک طرفه تحریک (یک تکانه عصبی می تواند تنها از غشای پیش سیناپسی به حالت پس سیناپسی منتقل شود).
تأخیر سیناپسی ، یعنی زمان مشخصی برای انتقال یک ضربه عصبی در سیناپس صرف می شود. (سرعت انتقال سیناپسی به طور متوسط \u200b\u200bبیش از 10 برابر سرعت انتشار ضربان عصبی از طریق عصب است. برای یک سیناپس شیمیایی معمولاً بین 0.2-0.5 میلی ثانیه است).
خستگی - کاهش تدریجی یا قطع کامل انتقال یک تکانه عصبی با تحریک عصبی طولانی مدت. اساس این پدیده از یک طرف کاهش ذخایر انتقال دهنده عصبی در ضخامتهای پیش از تحرک و از طرف دیگر کاهش حساسیت به گیرنده های پس سیناپسی به انتقال دهنده عصبی است.
حساسیت بالای سازندهای سیناپسی به داروها و سموم.
این آخرین ویژگی سیناپسها است که کل داروسازی داروهایی که بر فعالیت عملکرد سیناپسهای واقع در اندامها و بافتهای مختلف بدن تأثیر می گذارد ، مستقر است. باید تأکید کرد که موضوع عمل دارویی می تواند هر یک از مراحل انتقال سیناپسی باشد. به عنوان داروهایی که بر انتقال سیناپسی تأثیر می گذارند ، از آنالوگ های اگزوژن انتقال دهنده های عصبی ، پیش سازهای شیمیایی آنها و سایر مواد فعال بیولوژیکی که به هر طریق می توانند فعالیت عملکردی سیناپس را تغییر دهند استفاده می شود.
لازم به ذکر است که بسیاری از داروها دارای یک اما چند نکته کاربرد اثر در سطح سیناپس نیستند. به عنوان مثال ، داروی ضد افسردگی پیرلیندول نه تنها فعالیت آنزیم مونوآمین اکسیداز در شکاف سیناپسی را مهار می کند ، بلکه بازگرداندن نوراپی نفرین را توسط غشای پیش سیناپسی نیز مسدود می کند.
در رابطه با محلی سازی گیرنده به سیناپس ، می توان آنها را به پیش سیناپسی ، پس سیناپسی و خارج از سیناپسی تقسیم کرد. بعنوان مثال ، گیرنده های موجود در غشای سلولی پلاکت ها قرار دارند.
از نظر توپوگرافی سلولی (محل) ، گیرنده ها نیز می توانند با توجه به محل قرارگیری آنها بر روی ساختارهای سلولی به شرح زیر طبقه بندی شوند:
گیرنده های غشایی - گیرنده های واقع در غشای سیتوپلاسمی.
گیرنده های سیتوزولی - گیرنده های واقع در سازندهای داخل سلولی.
گیرنده های هسته ای - گیرنده های واقع در غشای هسته سلول.
همانطور که قبلاً نیز اشاره شد ، در نتیجه تعامل با گیرنده مواد بیولوژیکی درون زا یا داروها ، فعالیت عملکردی سلولهای هدف تغییر می کند. این فرایند می تواند به روش های مختلفی اجرا شود ، که برای انواع مختلف گیرنده ها کاملاً تعریف شده است. مطابق با این ، در حال حاضر چهار نوع گیرنده متمایز شده است که هرکدام دارای ویژگی های خاص خود هستند ، که اساساً با سایرین متفاوت است ، مکانیسم بواسطه آن سیگنالی از گیرنده ، آبشار واکنشهای بیوشیمیایی و / یا بیوفیزیکی را آغاز می کند که منجر به تغییر وضعیت عملکرد سلولهای هدف می شود.
سه نوع گیرنده اول روی غشای سلولی (سیتوپلاسمی) بومی سازی شده و نوع چهارم گیرنده ها شامل گیرنده های سیتوزولی و هسته ای است.
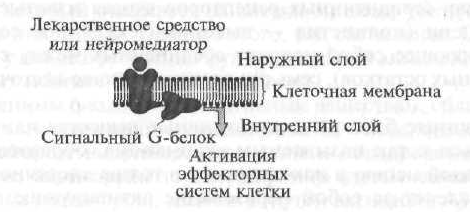
گیرنده های نوع I شامل گیرنده های سلولی (غشایی) هستند که تأثیرات خود را از طریق به اصطلاح پروتئین های G سیگنالینگ متوجه می شوند (شکل 1.7).
در مرحله اول ، یک ماده یا داروی بیولوژیکی فعال ، "بالا رفتن" به غشای سلولی ، گیرنده را "تشخیص" داده و با آن تعامل می کند ، پس از آن گیرنده یک پروتئین G ویژه سیگنال مستقر در سطح داخلی غشاء را فعال می کند. علاوه بر این ، پروتئین G فعال شده فعالیت عملکردی عنصر سازنده داخلی را تغییر می دهد ، که ، به عنوان یک قاعده ، آنزیم ها هستند. سپس عنصر مؤثر ، که آنزیمی است ، یک پیام رسان ثانویه یا یک پیام رسان ثانویه را فعال می کند ، که باعث ایجاد آبشار واکنشهای بیوشیمیایی می شود که فعالیت عملکردی سلولهای هدف را تغییر می دهد.
گیرنده های سلول نوع I ، یعنی گیرنده های کونژوگه شده با سیگنال پروتئین G از نظر ساختاری شبیه به یکدیگر هستند و در سازمان فضایی آنها مارپیچ (از مارپانت فرانسوی - مار ، توپ) ساختار هستند (شکل 1.8).
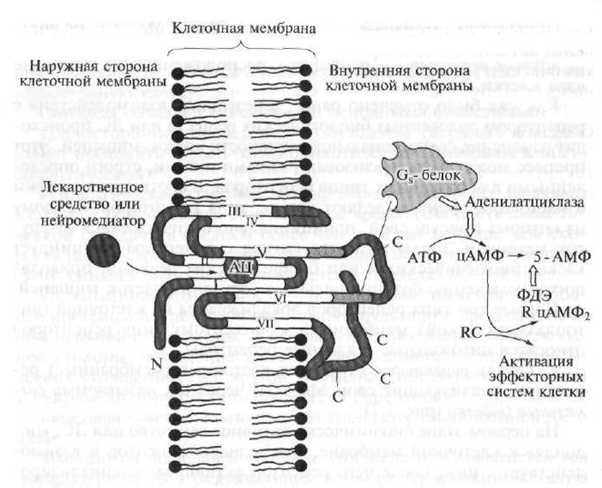
شکل 1.7 ساختار گیرنده نوع 1 (توضیح در متن)
شکل 1.8 نمایش شماتیک ساختار "مار"
گیرنده:
N قسمت پلی پپتیدی گیرنده واقع در بالای غشای سلولی است. C بخش پلی پپتیدی گیرنده واقع در زیر غشای سلولی است. AC - مرکز فعال گیرنده ای که دارو با آن تعامل دارد ؛ انتقال دهنده عصبی است. ATP - آدنوزین تری فسفات - پیام رسان ثانویه؛ cAMP - مونوفسفات آدنوزین حلقوی؛ 5-AMP - آدنوزین -5 "مونوفسفات ؛ PDE - فسفودی استراز ؛ R. RC - آنزیم وابسته به اردوگاه (پروتئین کیناز) با زیر واحد های نظارتی و کاتالیزوری (تسریع در واکنش) ؛ 1-VII - زنجیره های پلی پپتید گیرنده مارپیچ
گیرنده های مارپیچ شامل زنجیره های پلی پپتیدی پیچیده است (یک پلی پپتید یک ترکیب با وزن مولکولی بالا است ، که یک زنجیره مانده با اسید آمینه به هم پیوسته است) که هفت بار در غشای سلولی نفوذ می کند.
مواد یا داروهای بیولوژیکی فعال درون زا می توانند به اصطلاح "جیب" ایجاد شده توسط زنجیره پلی پپتید و در ضخامت غشای سلولی مستقر شوند ، که مستلزم تشکیل سیگنال فعال کننده است ، که به بخش هایی از زنجیره گیرنده واقع در سیتوپلاسم سلول منتقل می شود. پروتئین های سیگنال G با سی-
بخش های دزدی (داخل سلولی) کف و زنجیره پپتید. فعال کردن و راه\u200cاندازی یک آبشار از واکنشهای بیوشیمیایی که فعالیت عملکردی آن را در مورد سلول هدف تغییر می دهد ، یعنی شروع یک پاسخ دارویی اولیه.
در حال حاضر ، چندین نوع پروتئین سیگنال G شناخته شده است.
سیگنال G ، پروتئین ها. این پروتئین های سیگنالینگ ، به عنوان یک قاعده ، یک عنصر مؤثر را فعال می کنند - آنزیم آدنیلات سیکلاز ، که به نوبه خود باعث تحریک سنتز در سلول (از ATP) یک پیام رسان ثانویه - مونوفسفات آدنوزین حلقوی (cAMP) می شود. نقش بیولوژیکی cAMP به عنوان یک پیام رسان ثانویه بسیار مهم است. به عنوان مثال ، افزایش محتوای آن در سلولهای قلبی مستلزم افزایش فراوانی و قدرت انقباضات قلب است. علاوه بر این ، افزایش غلظت cAMP در سلولهای هدف مختلف باعث شل شدن عضلات صاف رگها و برونشها ، بسیج ذخایر انرژی (پوسیدگی کربوهیدرات ها در کبد) ، مهار توانایی تجمع پلاکت ها ، کاهش صدای میومتر (عضله رحم) و مثانه و غیره می شود.
تعدادی از انتقال دهنده های عصبی ، مانند آدرنالین (با فعال کردن گیرنده های β آدرنرژیک β) ، دوپامین (با فعال کردن گیرنده های D1-dopamine) ، آدنوزین (با فعال کردن گیرنده های آدنوزین A 2) ، متعلق به مواد فعال بیولوژیکی درون زا با توانایی فعال سازی پروتئین های سیگنالینگ Gs می باشد. هیستامین (با فعال سازی هیستامین G 2-رسپتوروپ) ، سروتونین (با فعال شدن گیرنده های 5-HT 4 سروتونین) ، و همچنین تعدادی از هورمون ها ، به عنوان مثال ، وازوپرسین (با تحریک گیرنده های V--vasopressin) و غیره
سیگنال G i پروتئین ها. بر خلاف پروتئین های سیگنال G ، فعال سازی پروتئین های سیگنال G i باعث تحریک نمی شود بلکه فعالیت عنصر مؤثر ، آنزیم آدنیلات سیکلاز را مهار می کند ، که منجر به کاهش غلظت cAMP در سلولهای هدف پیام رسان ثانویه می شود. کاهش محتوای cAMP در سلولهای هدف باعث کاهش انقباضات قلبی ، افزایش لحن عروق خونی و برونش ها می شود ، یعنی. اثر خلاف افزایش محتوای cAMP در سلولهای هدف. علاوه بر این ، تعدادی از پروتئین های G سیگنالینگ در تنظیم فعالیت عملکردی کانال های یونی Ca2+ و K + transmembrane نقش دارند.
تعدادی از انتقال دهنده های عصبی ، به عنوان مثال ، آدرنالین و نوراپی نفرین (با فعال کردن یک 2-گیرنده گیرنده) ، دوپامین (با فعال کردن گیرنده های D2 - دوپامین) ، آدنوزین (با فعال سازی A 1) متعلق به مواد بیولوژیکی فعال درون زا هستند که قادر به فعال شدن سیگنال G پروتئین ها هستند. گیرنده های آدنوزین) ، استیل کولین (با فعال سازی گیرنده های موسکارینی M2 و M4) و غیره
پروتئین G ^ سیگنال. این پروتئین های سیگنالینگ در فعال سازی یکی دیگر از مؤلفه های مؤثر سلولهای هدف ، آنزیم فسفوریلاز C ، که به نوبه خود باعث تحریک تشکیل پیام رسانهای ثانویه ، دیاسیل گلیسرول (DAG) و اینوزیتول-1،4،5-تری فسفات (ITP) در سلولهای هدف می شود. اولین آنها (DAG) با غشای سلولی همراه است و واکنشهای بیوشیمیایی در تنظیم وضعیت انقباض ، رشد و تقسیم سلولی و ترشح هورمونهای خاص توسط سلولهای هدف را آغاز می کند. تحت تأثیر آنزیم فسفولیپاز A 2 ، DAG می تواند به اسید آراشیدونیک متابولیزه شود ، که در سنتز مواد بیولوژیکی فعال مانند ایکوزانوئیدها - پروستاگلاندینی ، پروستاسیلین ها ، ترومبوکسان ها ، لکوترین ها متابولیزه می شود (نگاه کنید به T. I ، ص 478).
پیام رسان ثانویه ، ITF ، بر روی غشای سلولی ثابت نشده است و به داخل محیط داخل سلولی (سیتوزول) منتقل می شود ، جایی که آغاز آزاد سازی یون های Ca 2+ از انبارهای سلولی ، یعنی. انتقال یون های Ca 2+ غیرفعال به شکل فعال را ترویج می کند.
بسیاری از محققان یون های Ca 2+ را به عنوان یک پیام رسان سوم یا واسطه در نظر می گیرند. این در شرایطی است که نقش یون های Ca 2+ در تنظیم فعالیت عملکردی سلول ها بسیار مهم است. یون های Ca 2+ می توانند از طریق کانالهای یونی ویژه ترانسمیبرون وارد محیط شوند و یا از انبارهای سلول آزاد شوند. انبار اصلی (محل تجمع یون های Ca2+ غیرفعال) در سلول ، آندوپلاسمی ، یا سارکوپلاسمی ، رتیکول (reticulum sarcoplasmaticunr ، مترادف: شبکه آندوپلاسمیک - اندامکول داخل سلولی) است که سیستم توبولها و مخازن واقع در سیتوپلاسم ، محدود در غشاء است. حمل و نقل مواد در سیتوپلاسم). یون های Ca2 + رایگان (فعال) که از شبکه سارکوپلاسمی به سیتوپلاسم منتقل می شوند با برخی پروتئین های حاوی Ca 2+ ارتباط دارند که مهمترین آنها کالمودولین است. کمپلکسهای "کالمودولین-های Ca 2" و / یا مجموعه های یون های Ca2 با سایر پروتئین های اتصال کلسیم باعث ایجاد آبشار واکنشهای بیوشیمیایی در سلول می شوند. در نتیجه ، بسته به اندامهای هدف که در این فرآیند اتفاق می افتد ، افزایش عملکرد انقباضی میوکارد و عضله اسکلتی ، افزایش لحن عضلات صاف رگ های خونی ، برونش ها و رحم ، افزایش فعالیت ترشحی بافت غده ، تحریک رهایی از انتقال دهنده های عصبی از انتهای عصب و غیره آغاز می شود. . همچنین ثابت شده است که یون های Ca 2+ توانایی افزایش فعالیت آنزیم های درگیر در پروتئین ، کربوهیدرات و متابولیسم چربی را دارند.
علاوه بر رابطه مستقیم بین پیام رسان های ثانویه - DAG و ITF و بنابراین پروتئین های سیگنال G q ، یون های Ca 2+ در شرایط فیزیولوژیکی تعامل نسبتاً پیچیده ای با پیام رسان ثانویه cAMP دارند ، فعالیت آنها با سیگنال G و پروتئین G من تنظیم می شود. بنابراین ، نشان داده شده است که یون های Ca2+ رایگان که وارد سیتوپلاسم یک سلول عصبی از طریق سیستم کالمودولین-Ca 2+ می شوند ، باعث کاهش محتوای iAMP در سلول می شوند. در عین حال ، برای حفظ حالت باز کانالهای یون کلسیم در سلول ، غلظت بالای cAMP لازم است ، یعنی کاهش محتوای cAMP آغاز شده توسط مجتمع کالمودولین-Ca 2 + مستلزم قطع مصرف یون های Ca2 رایگان در سیتوپلاسم است. از طرف دیگر ، شواهدی وجود دارد که نشان می دهد پیام رسان ثانویه cAMP جذب یون های Ca2+ رایگان توسط رتیکولوم سارکوپلاسمی را تقویت می کند ، یعنی. انتقال یون های Ca 2+ از فرم آزاد و فعال به فرم محدود و غیرفعال را ترویج می کند.
در نتیجه افزایش محتوای پیام رسانهای ثانویه در سلولهای هدف - DAG و ITF - میزان تون عضلات صاف افزایش می یابد ، ترشح غدد افزایش می یابد ، آزاد شدن انتقال دهنده های عصبی از انتهای پیش سیناپی تسهیل می شود ، توانایی تجمع پلاکت ها و غیره فراهم می شود.
مواد فعال بیولوژیکی درون زا با توانایی فعال کردن پروتئین های سیگنال C q شامل انتقال دهنده های عصبی مانند نوراپی نفرین (با فعال کردن گیرنده های 1-adrenoreceptors) ، استیل کولین (با فعال کردن گیرنده های موسکارینی M1 و M3) ، سروتونین (به دلیل فعال شدن سروتونین گیرنده های 5-HT 2a) ، هیستامین (به دلیل فعال شدن گیرنده های هیستامین H 1) و همچنین سایر مواد فعال بیولوژیکی درون زا ، به عنوان مثال ، برادی کینین و آنژیوتانسین.
در حال حاضر ، علاوه بر سیگنال ذکر شده پروتئین های G (G ، G | ، G q) ، پروتئین های سیگنال دیگری نیز شناسایی شده اند - G s ، G i ، G q ، نقش فیزیولوژیکی آن هنوز کاملاً مشخص نیست. اما در عین حال ، شواهدی وجود دارد که ، به عنوان مثال ، پروتئین سیگنال C o در تنظیم فعالیت عملکردی کانالهای یونی transmembrane نقش دارد.
واحد عملکردی گیرنده های نوع 11 پروتئین (آنزیم) قابل عبور از ضخامت غشای سلول است. گیرنده خود شامل دو قطعه یکسان است که به آنها مونومر گفته می شود. مونومرها در فاصله ای ناچیز از یکدیگر قرار دارند و خود مونومر از دو زیر واحد عملکردی تشکیل شده است - حوزه ها ، که توسط یک قطعه پلی پپتیدی که با هم تلاقی غشاء لایه لیپیدی را تشکیل می دهد ، متصل شده است (شکل 1.9). a- زیر واحد مونومر بیرون از سطح بیرونی غشای بیرون زده و وظیفه اتصال گیرنده به مواد فعال بیولوژیکی را بر عهده دارد ، و زیر واحد P در سیتوپلاسم سلول غوطه ور است.

شکل 1.9 ساختار گیرنده نوع II (توضیح در متن): 1 - زیرواحد مونومر. 2 - β- زیر واحد مونومر
پس از اتصال ماده بیولوژیکی فعال به زیرواحد α گیرنده ، گیرنده از حالت غیرفعال مونومر به حالت دیمری فعال تغییر می یابد که در آن دو مونومر در صفحه غشاء ترکیب می شوند (شکل 1.9 را ببینید). در این حالت ، فعالیت آنزیمی سیتوپلاسمی β زیر واحد گیرنده تحریک می شود ، در نتیجه ، آبشار واکنشهای بیوشیمیایی که حالت عملکردی آن را تغییر می دهند در سلول هدف راه اندازی می شود.
به عنوان یک آنزیم transmembrane که گیرنده را تشکیل می دهد ، به عنوان یک قاعده ، از آنزیمی هایی مانند تیروزین کیناز یا گانیلات سیکلاز استفاده می شود.
نمونه ای از گیرنده تیروزین کیناز گیرنده های انسولین است (به تی 1 ، صفحه 435 مراجعه کنید).
مسیر انتقال سیگنال گانیلات سیکلاز با تعامل یک زیر واحد گیرنده با یک ماده فعال بیولوژیکی درون زا مثلاً با فاکتور ناتریورتیک دهلیزی (ANF) شروع می شود ، که یک ماده فعال بیولوژیکی است که توسط سلولهای دهلیزی ترشح می شود و در تنظیم انقباضات قلبی نقش دارد. در نتیجه این تعامل ، تغییری در پیکربندی گیرنده رخ می دهد ، که در ترکیب مونومرهای آن به یک دیمر تشکیل شده است. این فرآیند قسمت آنزیمی گیرنده واقع در β زیرواحد سیتوزولی آن را فعال می کند ، یعنی. آنزیم گوانلات سیکلاز ، که به نوبه خود باعث افزایش غلظت پیام دهنده ثانویه حلقوی گوانیدین 3،5 "مونوفسفات (cGMP) در سلول هدف می شود. افزایش غلظت cGMP در سلولهای هدف باعث ایجاد آبشار از واکنشهای بیوشیمیایی می شود که حالت عملکردی آنها را تغییر می دهد ، به عنوان مثال ، آرامش سلولهای عضله صاف. کشتی ها
گیرنده های نوع III شامل گیرنده هایی هستند که تحت تأثیر مواد بیولوژیکی فعال درون زا - انتقال دهنده های عصبی ، از عبور یون های مربوطه از طریق غشای سلولی اطمینان می کنند ، که منجر به تغییر در بار الکتریکی (ممبران) آن (پتانسیل) می شود.
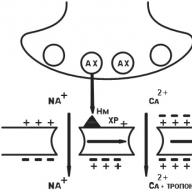
در ساختار آنها ، گیرنده های III tina نشان دهنده ی كانالی هستند كه در لایه چربی غشای سلولی نفوذ می كنند و توسط چندین واحد لوله یك لوله ساخته می شوند (شکل 1.10). به عنوان مثال ، گیرنده نیکوتین (H) یک کانال با قطر 8 نانومتر است که توسط پنج زیر واحد پلی پپتیدی (a - دو ، β ، γ ، d) تشکیل می شود (شکل 1.10 را ببینید). هنگامی که استیل کولین انتقال دهنده عصبی با بخشی (دامنه) بیرون زده از سطح غشای سلولی - زیر واحد α گیرنده - در تعامل باشد و ساختار آن تغییر کند و یک کانال مرکزی باز شود ، از طریق آن یون های Na + مطابق با شیب غلظت وارد سلول مورد نظر می شوند که این امر مستلزم تغییر در عملکرد آن است. فعالیت علاوه بر گیرنده های H- کولینرژیک ، گیرنده های اسید گاما آمینوبوتیریک و اسیدهای آمینه تحریکی متعلق به گیرنده های نوع III هستند.
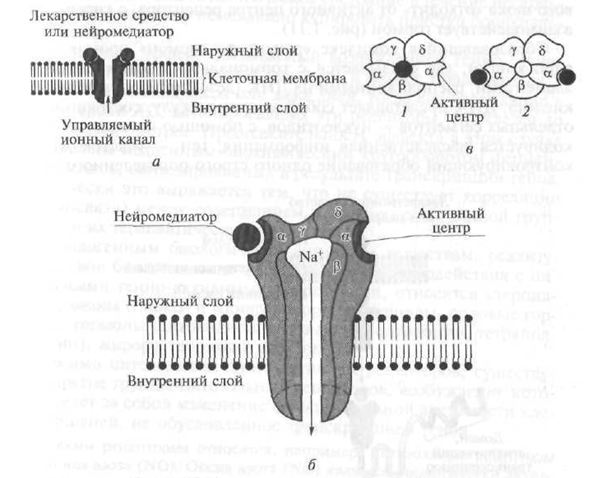
شکل 1.10 نمودار ساختار گیرنده نوع 111:
a یک نمودار مدار است. b - کانال یونی transmembrane (در متن)؛ ج - کانال یونی transmembrane (نمای بالا)؛ / - کانال در حالت غیرفعال (بسته) 2 - کانال در حالت فعال (باز)؛ یک زیر واحد های پلی پپتیدی β ، γ ، d - کانال
گیرنده های نوع IV شامل گیرنده های درون سلولی و هسته ای هستند. مواد فعال بیولوژیکی متقابل با این نوع گیرنده ها ترکیبات لیپوفیلیک (به راحتی محلول در چربی ها) هستند ، بنابراین ، آنها به راحتی در غشای سلولی نفوذ کرده و به گیرنده های داخل سلول خود می رسند. گیرنده های داخل سلولی شامل گیرنده های هورمون ها و سایر مواد فعال بیولوژیکی هستند.
مکانیسم تعامل هورمون ها با گیرنده های داخل سلولی کاملاً پیچیده است ، با این حال می توان آن را به صورت شماتیک به شرح زیر نشان داد. با توجه به ساختار ، گیرنده داخل سلولی هورمون ها یک پلی پپتیدی است که از چندین واحد عملکردی - حوزه تشکیل شده است. در صورت عدم وجود هورمون ، گیرنده غیرفعال است زیرا مرکز فعال آن توسط یک پروتئین تخصصی مسدود شده است - به اصطلاح پروتئین شوک حرارتی. در مواردی که هورمون "به" گیرنده نزدیک می شود ، پروتئین شوک حرارتی "از مرکز فعال گیرنده" خارج می شود "، که با آن هورمون ارتباط برقرار می کند" ("شکل 1.11).
مجتمع گیرنده هورمون گیرنده به هسته هسته سلول نفوذ می کند ، جایی که به عناصر حساس به هورمون مستقر در DNA (اسید دئوکسییریبونوکلئیک ؛ DNA متصل می شود) ، ماکرومولکولی است که از بخش های جداگانه - نوکلئوتیدها تشکیل شده و به کمک آن اطلاعات ارثی در ژن ها رمزگذاری می شود ؛ ژن - قطعه ای از DNA که تشکیل پروتئین کاملاً تعریف شده را کنترل می کند
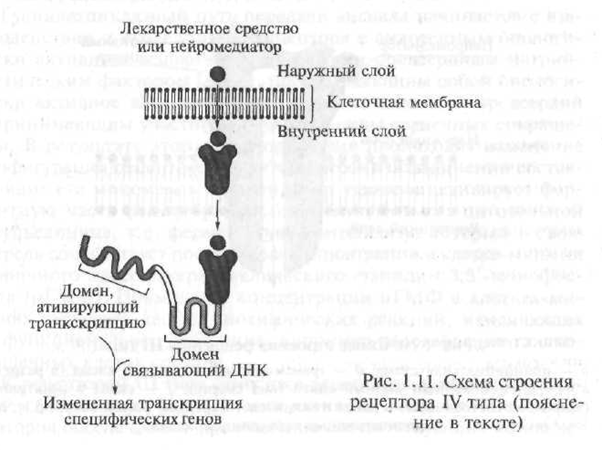
شکل 1.11 طرح ساختار گیرنده نوع IV (توضیح در متن)
کا) در نتیجه این تعامل ، فرآیند رونویسی ژن آغاز می شود - فرایند انتقال اطلاعات موجود در کد ژنتیکی از مولکول DNA به مولکول RNA اطلاعات (mRNA ، syn: matrix RNA - mRNA). رونویسی اولین قدم در شکل گیری پروتئین ها در یک سلول است. mRNA حاصل. هسته سلول را ترک می کند و به سمت ریبوزوم ها حرکت می کند - اندامک های درون سلول که مسئول سنتز پروتئین در سلول هستند. در ادبیات پزشکی خاص ، به گیرنده هایی که فعال شدن آنها باعث فرآیند رونویسی ژن ها می شود ، به گیرنده های فعال ژنتیکی گفته می شود.
به عنوان یک قاعده ، پاسخ سلولهای هدف به تحریک گیرنده های فعال ژن نسبتاً آرام ایجاد می شود ، که از اهمیت بالینی بسیار مهمی برخوردار است.
اول ، پاسخ سلولهای هدف به موقع به تأخیر می افتد ، زیرا به سنتز پروتئین های جدید نیاز دارد ، که معمولاً 20-30 دقیقه طول می کشد ، یعنی. هورمون ها ، گیرنده های فعال کننده نوع IV ، قادر به تغییر وضعیت پاتولوژیک در طی چند دقیقه نیستند ، به عنوان مثال ، بلافاصله حمله آسم برونشی را متوقف کنید.
ثانیا ، اثر ناشی از تحریک گیرنده های فعال ژن بسیار طولانی است و می تواند چندین ساعت یا حتی چند روز طول بکشد ، در حالی که محتوای موجود در پلاسمای خون داروهای فعال کننده این گیرنده ها بسیار سریعتر به صفر می رسد. مدت اثر در این مورد به دلیل گردش بیوشیمیایی کند آنزیم ها و پروتئین های سنتز شده در نتیجه رونویسی ژن است. از نظر بالینی این امر با این واقعیت بیان می شود که هیچ ارتباطی بین رابطه پلاسمائی یک گروه خاص از داروها و تأثیر درمانی آنها وجود ندارد.
مواد فعال بیولوژیکی درون زا که اثرات بیولوژیکی خود را از طریق تعامل با گیرنده های فعال ژن سیتوزولی درک می کنند شامل هورمونهای استروئیدی (گلوکو و کانی های کانی ، کوره های هورمونهای جنسی) ، هورمونهای تیروئید (تریودوتیرونین ، تتراودوتیرونین) و ویتامین D محلول در چربی است.
علاوه بر گیرنده های فعال ژن سیتوزولی ، گروه های دیگری از گیرنده های سیتوزولی نیز وجود دارند که تحریک آنها مستلزم تغییر در عملکرد عملکردی سلولهای هدف است ، نه به دلیل رونویسی ژنها.
چنین گیرنده هایی شامل ، گیرنده های سیتوزولی برای اکسید نیتریک (N0) هستند. اکسید نیتریک (N0) یک ماده فعال بیولوژیکی است که در اندوتلیوم عروقی تشکیل می شود. به عنوان یک ماده فعال بیولوژیکی درون زا ، اکسید نیتریک برای اولین بار در سال 1987 توسط فیزیولوژیست آمریکایی R.F. Furchgott از بنادر خرگوش جدا شد و "عامل آرامش بخش اندوتلیال - ORF" نامیده شد. اکسید نیتریک یک ترکیب لیپوفیلی است که به راحتی در غشای سلولی نفوذ می کند ، جایی که با گیرنده های سیتوزولی خاص آن تعامل دارد و این امر مستلزم فعال سازی آنزیم گوانلات سیکلاز است. دومی ، به نوبه خود ، سنتز پیام رسان ثانویه cGMP را تحریک می کند ، که باعث ایجاد آبشار واکنشهای بیوشیمیایی داخل سلولی و منجر به شل شدن سلولهای هدف ، سلولهای عضلانی صاف عروقی می شود.
بنابراین ، در حال حاضر ، چهار مکانیسم اصلی و بر این اساس ، انواع گیرنده IV از هم متمایز می شوند ، به دلیل تعامل با آنها مواد فعال بیولوژیکی درون زا و / یا آنالوگ های مصنوعی آنها ، یعنی. داروها می توانند بر عملکرد سلولهای هدف تأثیر بگذارند.
با این حال ، این بدان معنی نیست که تعداد گیرنده های شناخته شده برای مواد فعال بیولوژیکی به 4 محدود می شوند. این در شرایطی است که از طریق همان مکانیسم اساسی عمل ، تعداد بسیار زیادی از مواد بیولوژیکی درون زا از ساختارهای مختلف شیمیایی می توانند بر فعالیت عملکردی سلولها تأثیر بگذارند. به عنوان مثال ، انتقال دهنده های عصبی نوراپی نفرین و هیستامین ، که در ساختار شیمیایی آنها متفاوت است و بنابراین ، در گیرنده هایی که با آنها تعامل دارند ، یک سیگنال تحریکی را با همان مکانیسم بنیادی - تحریک فعالیت پروتئین های سیگنال G ، یعنی سلول تحریک می کنند. هر دو با گیرنده های نوع I در تعامل هستند.
بنابراین ، تمام گیرنده های شناخته شده در حال حاضر نه تنها بر اساس ویژگی های انتقال سیگنال به ساختارهای داخل سلول سلولهای هدف طبقه بندی می شوند ، بلکه همچنین بر روی نام آنها از مواد بیولوژیکی فعال درون زا که با آنها به طور خاص تعامل دارند ، طبقه بندی می شوند.
لازم به ذکر است که گیرنده ها مدت ها قبل از شناخته شدن مکانیسم های انتقال سیگنال به سلولهای هدف ، نام خود را با در نظر گرفتن نام مواد فعال بیولوژیکی درون زا که با آنها ارتباط برقرار می کنند ، به دست آوردند.
مواد فعال بیولوژیکی درون زا که اثرات آنها را از طریق تعامل با گیرنده های خاص خود متوجه می شوند شامل انتقال دهنده های عصبی (استیل کولین ، نوراپی نفرین ، دوپامین ، هیستامین ، سروتونین و غیره) ، هورمون ها ، مواد بیولوژیکی فعال با منشاء بافت - اتوکائیدها (پروستاگلاندین ها ، ترومبوکسان ها ، لکوترین ها ، برادی کینین ، آنژیوتانسین و غیره). در ادبیات پزشکی ویژه ، همه این مواد معمولاً تحت عنوان "لیگاند" (از لاتین لیگو - برای اتصال ، یعنی ماده ای که قادر به اتصال به گیرنده است) ترکیب می شوند.
بنابراین ، گیرنده ها نام خود را از نام لیگاند های خاص خود دریافت می کنند. به عنوان مثال ، گیرنده های انتقال دهنده عصبی دوپامین به نام های دوپامین ، هورمون انسولین - انسولین ، لکوترین ها آگوکائید - لکوترین و غیره گفته می شود.
در آینده ، در متن کتاب درسی ، به منظور جلوگیری از سردرگمی در اصطلاحات در هنگام ساز و کار انتقال سیگنال از گیرنده به سازندهای درون سلولی ، از اصطلاح "نوع گیرنده" استفاده می شود و هنگامی که از نام گیرنده به دلیل تعامل لیگاند با آن استفاده می شود ، استفاده می شود. از اصطلاح "گونه های گیرنده" استفاده خواهد شد.
به عنوان یک قاعده ، بسیاری از گیرنده های همان گونه به چندین زیرگروه تقسیم می شوند ، به عنوان مثال ، گیرنده های آدرنرژیک به گیرنده های a- و β- آدرنرژیک ، گیرنده های کولینرژیک به گیرنده های M- و N- کولینرژیک و غیره تقسیم می شوند. در بیشتر موارد ، زیر گونه ها به گروه های کوچکتر نیز تقسیم می شوند: گیرنده های گیرنده β1- و β2 ، گیرنده های N-N و N- کلینرژیک و غیره:
شناسایی زیرگونه گیرنده ها و مطالعه مکانیسم هایی که مواد بیولوژیکی فعال درون زا با آنها در تعامل هستند ، برای داروسازی مدرن بسیار مهم است ، زیرا امکان ایجاد داروهایی را می دهد که با یک زیرگونه کاملاً تعریف شده گیرنده در تعامل هستند. بنابراین ، به عنوان مثال ، تقسیم گیرنده های β- آدرنرژیک به β 1 (عمدتا در محل غشای سلول های قلب بومی سازی شده) و β2 (بومی سازی شده ، به عنوان مثال ، روی غشای سلول های عضلات صاف برونش ها) ، امکان ایجاد داروهایی را فراهم می کند که بطور انتخابی بر روی عضله قلب تأثیر می گذارند (β 1-آدرنوستیمولانتها ) داروی nonachlazine ، و انتخابی بر عضلات صاف برونشها تأثیر می گذارد (β2-adrenostimulants) - سالبوتامول دارو ، و غیره.
لازم به ذکر است که اندام ها و بافت های بدن حاوی تعداد ثابت گیرنده و / یا زیرگونه آنها نیست ، یعنی. متغیر است هم فرایندهای آسیب شناختی و هم داروها می توانند تعداد گیرنده های موجود در اندام را تغییر دهند.
به عنوان مثال بیماری عروق کرونر قلب با افزایش تعداد گیرنده های 3-آدرنرژیک در عضله قلب همراه است و در بیمارانی که فشار خون بالا دارند ، تعداد گیرنده های a- و β- آدرنرژیک افزایش می یابد. ایمی پرامین ضد افسردگی با مصرف طولانی مدت ، میزان گیرنده های β آدرنرژیک در بافت مغز را کاهش می دهد. نمونه های بسیار مثالی از این دست وجود دارد.
قرابت انتقال دهنده های عصبی درون زا (تولید شده در بدن) یا داروها برای گیرنده ها با اصطلاح "میل" مشخص می شود و سرعت و قدرت اتصال آنها به گیرنده ها با اصطلاح "میل" مشخص می شود.
به طور طبیعی ، تعامل داروها با گیرنده به خودی خود پایان نمی یابد ، بلکه باید به برخی تغییرات فعالیت اندام ها یا بافت های بدن منجر شود.
چنین تغییر یا واکنشی متناسب با اهمیت عملکردی این گیرنده ، فعالیت داخلی داروها نامیده می شود.
داروهایی با فعالیت ذاتی و میل به گیرنده آگونیست ها هستند ، یعنی مانند مواد بیولوژیکی فعال درون زا عمل کنید.
به عنوان مثال ، فنیل افرین ، یک محرک گیرنده های آدرنرژیک ، در شریانی مشابه با نوراپی نفرین عصبی انتقال دهنده عصبی ، یعنی تأثیر دارد. او آگونیست گیرنده های آدرنرژیک است. در ادبیات پزشکی خاص ، علاوه بر اصطلاح "آگونیست" ، اصطلاح "محرک گیرنده" یا "mimetic" گاهی اوقات نیز به کار می رود ، به عنوان مثال ، آدرنومیمی ، یعنی. داروهایی که گیرنده های آدرنرژیک را تحریک می کنند.
داروهایی که میل به گیرنده دارند اما آگونیست های برون زا و درون زا را از تعامل با گیرنده مهار می کنند ، آنتاگونیست نامیده می شوند.
به عنوان مثال ، مسدودکننده گیرنده گیرنده آتروپین M- کولینرژیک با تعامل گیرنده عصبی استیل کولین M- کولینرژیک تداخل می کند. مسدود کننده (گیرنده های J- آدرنرژیک پروپرانولول ، مسدود کردن β2-آدرنرژی ساختار ریه ها ، مانع از اثر تحریک کننده β2-آدنروستیمولاتور سالبوتامول بر روی آنها ، یعنی آتروپین و پروپرانولول آنتاگونیست گیرنده های مربوطه می باشد.
در ادبیات تخصصی پزشکی ، علاوه بر اصطلاح "آنتاگونیست" ، اصطلاح "مسدود کننده های گیرنده" یا "لیتیک" ، به عنوان مثال ضد کولینرژیک ، یعنی. داروهایی که گیرنده های کولینرژیک را مسدود می کنند.
به طور شماتیک ، تعامل گیرنده ها با آگونیست ها و آنتاگونیست ها در شکل نشان داده شده است. 1.12
آگونیست ها می توانند هم مستقیم و هم غیرمستقیم عمل کنند ، یعنی عمل غیر مستقیم

شکل 1.12 طرح تعامل گیرنده با آگونیست (a) و آنتاگونیست (6) با گیرنده (توضیح در متن)
به عنوان مثال ، آگونیست گیرنده مواد افیونی - مرفین دارو - اثرات آن را با تحریک مستقیم (mu) p- ، (kappa) k- و (delta) گیرنده های 8 افیونی ، یعنی تحقق می یابد. یک اثر تحریک کننده مستقیم بر روی آنها اعمال می کند ، در حالی که افدرین سمپاتومیمیک با تحریک غیر مستقیم یا غیرمستقیم گیرنده های a- و β- آدرنرژیک اثرات سلولی خود را تحقق می بخشد: این امر به جابجایی نوراپی نفرین عصبی انتقال دهنده عصبی از انتهای عصب پیش سیناپسی ، بازگرداندن مجدد آن توسط انتهای عصب ، و افزایش حساسیت- گیرنده های آدرنرژیک به نوراپی نفرین و اپی نفرین و تحریک ترشح آدرنالین از قشر آدرنال را تحریک می کند. بنابراین ، افدرین اثرات دارویی خود را نه به طور مستقیم ، بلکه از طریق یک انتقال دهنده عصبی ، می فهمد ، در حالی که به طور مستقیم با گیرنده تعامل ندارد.
آنتاگونیست ها و همچنین آگونیست های گیرنده می توانند اثرات دارویی خود را بطور مستقیم یا غیرمستقیم متوجه شوند و گیرنده های مربوطه را مسدود کنند.
به عنوان مثال ، مسدود کننده گیرنده هیستامین معده H 3 ، رانیتیدین ، \u200b\u200bبه طور مستقیم با گیرنده های هیستامین معده H2 ارتباط برقرار می کند ، ترشح اساسی اسید هیدروکلریک را مهار می کند ، تعامل با گیرنده عصبی هیستامین هیستامین درون زا ، مهار می کند. به طور مستقیم گیرنده را مسدود می کند ، در حالی که رزرپین سمپاتولیتیک ، وارد بدن می شود ، مانند افدرین سمپاتیوماتیک نورپین افرین نوروترانسمیتر را از انتهای پیش سیناپسی خارج می کند ، در حالی که همزمان سنتز "بخش های" جدید نوراپی نفرین ، که منجر به تجمع سریع مدیون می شود ، مسدود می شود. در نتیجه ، وضعیتی ایجاد می شود که گیرنده های آدرنرژیک عروق به دلیل عدم تحریک کننده فیزیولوژیکی نورآدرناپین ، کار نمی کنند. می توان گفت که رزرپین سمپاتولیتیک باعث انسداد عملکردی و غیرمستقیم گیرنده های یک آدرنرژیک می شود.
بعضی از داروها خواص آگونیست ها و آنتاگونیست ها را ترکیب می کنند ، یعنی. تحت شرایط خاص ، همان گیرنده ها هیجان زده یا مسدود می شوند. در مواردی که ماده تحریک کننده در اثر دارویی دارو غالب باشد ، می گویند دارو آگونیست جزئی یا جزئی است.
مکانیسم عمل آگونیست های جزئی یا جزئی این است که این داروها ، با نزدیک شدن به گیرنده ، روی آن ثابت شوند. با این حال ، به دلیل ویژگی های ساختار شیمیایی آنها ، آنها را به شکلی هیجان زده می کنند که باعث می شوند نه یک واکنش کامل ، بلکه فقط یک واکنش تحریک کننده جزئی باشد. در همان زمان ، آنها در تعامل سایر داروها و / یا انتقال دهنده عصبی مربوطه با این گیرنده دخالت می کنند.
به عنوان مثال ، نالورفین ، که برای درمان مصرف بیش از حد از داروهای ضد درد مواد مخدر استفاده می شود ، آگونیست جزئی است. نالورفین ، "بالا رفتن" به گیرنده مواد افیونی ، مورفین را از اتصال آن جدا می کند ، یعنی اثر آگونیستی مورفین را بر روی گیرنده متوقف می کند. تماس با گیرنده ، نالورفین تا حدی تأثیر تحریک کننده ای بر روی آن دارد که قادر به سرکوب فعالیت مرکز تنفسی نیست ، یعنی. دارای یک اثر آگونیستی جزئی بر روی گیرنده مواد افیونی است ، به موازات آن ، آن را برای مورفین مسدود می کند ، در نتیجه با توجه به مورفین اثر متضاد به نمایش می گذارد. ضرر نالورفین این است که در صورت مصرف بیش از حد ، اثر تحریک کننده آن بر روی گیرنده تقویت شده و باعث مهار مجاری تنفسی و همچنین مرفین می شود.
در مواردی که اثر مسدودکننده در اثر دارویی داروها حاکم باشد ، می گویند که این دارو با فعالیت داخلی یا خاص خود یک آنتاگونیست است.
ویژگی های مکانیسم عملکرد آنتاگونیست ها با فعالیت خود را می توان به عنوان مثال β 1-adrenoblocker acebutalol در نظر گرفت. Acebutalol متعلق به گروه مسدود کننده های آدرنرژیک β 1 با فعالیت سمپاتومیمی داخلی آنها ، یعنی. ترکیبی از خواص مسدود کننده (لیتیک) و یک محرک (تقلید) از β 1-آدرنرژیک است.
مکانیسم عمل آکوبوتالول به دلیل ویژگیهای ساختار شیمیایی آن است ، به همین دلیل دارو از یک طرف ، میوکارد β 1-آدرنرفتورها را مسدود می کند ، و از سوی دیگر ، فعالیت آنها را تحریک می کند ، از این طریق اثر فیزیولوژیکی واسطه ها - کاتکول آمین ها را شبیه سازی می کند. 170)
کلیه داروها با توجه به خصوصیات عملکرد آنها می توانند به دو گروه بزرگ تقسیم شوند - با اثرات خاص و غیر اختصاصی.
داروهایی که خاصیت غیر اختصاصی دارند شامل داروهایی است که دامنه وسیعی از فعالیت دارویی و نقاط مختلف استفاده از اثرات دارند. این گروه شامل ویتامین ها ، محرک های بیوژنیک ، آنتی اکسیدان ها ، سازگاری ها و غیره است.
داروهایی با اثر خاص شامل داروهایی هستند که با خاصیت آگونیستها یا آنتاگونیست های گیرنده های مربوطه ، مشخص می شوند. اساس مکانیسم عمل آنها توانایی تعامل با گیرنده های خاص برای آنها است.
با این حال ، باید توجه داشت که اثر دارویی داروهایی که تأثیر خاصی روی هر گیرنده دارند ، همیشه تخصص خاصی ندارند ، به عنوان مثال کاهش ضربان قلب (HR) یا سرکوب ترشح آب معده. اثر دارویی کل بسیاری از داروها بستگی به تعداد و در چه اعضای و بافتهای سازندهای گیرنده اختصاصی بدن برای این دارو دارد.
به عنوان مثال ، گیرنده های β آدرنرژیک در عضله قلب ، دیواره عروق ، برونش ، رحم ، بافت چربی ، عضله اسکلتی و غیره قرار دارند. در نتیجه این ، داروهایی که گیرنده های β- آدرنرژیک را تحریک می کنند ، تا حدودی باعث افزایش قدرت و فرکانس انقباضات قلبی ، گسترش رگ های خونی ، برونش ها و صدای پایین تر رحم می شوند ، یعنی تأثیر سیستمیکی روی بدن دارند.
تأثیر خاص داروها می تواند انتخابی یا انتخابی باشد و بر همین اساس غیر انتخابی یا غیر انتخابی باشد. انتخاب داروها با اینكه آیا همه زیر گونه های گیرنده ها یا زیرگونه های خاص آنها را تحت تأثیر قرار می دهد ، تعیین می شود.
به عنوان مثال ، مسدود کننده β- آدرنرژیک پروپرانولول غیر انتخابی هر دو گیرنده β1-آدرنرژیک واقع در بافت میوکارد و β 2-آدرنرژیک واقع در ، به ویژه ، در بافت ریه ، به عنوان مثال ، یک اثر غیر انتخابی (غیر انتخابی) در هر دو گونه گیرنده β آدرنرژیک دارد. . در حالی که آتنولول مسدودکننده β 1 آدرنرژیک انتخابی در دوزهای درمانی تنها گیرنده های آدرنرژیک β1 میوکارد را مسدود می کند ، یعنی. اثر انتخابی (انتخابی) بر روی گیرنده β آدرنرژیک خاص دارد.
شفاف سازی نقش فیزیولوژیکی و بومی سازی گونه های گیرنده امکان ایجاد داروهای بسیار مؤثر را دارد که به صورت انتخابی در زیر گونه های مختلف گیرنده ها عمل می کنند.
به عنوان مثال ، داروهایی که گیرنده های β-آدرنرژیک β 1 را مسدود می کنند ، به طور گسترده ای در درمانگاه برای درمان بیماری عروق کرونر قلب ، فشار خون شریانی و داروهایی که گیرنده های β 2 آدرنرژیک را تحریک می کنند ، کاربردهای خود را برای درمان آسم برونش یافتند. داروهایی که گیرنده های هیستامین H1 را مسدود می کنند ، ترجیحاً در عمل بالینی برای جلوگیری و / یا دستگیری تجویز می شوند واکنشهای آلرژیکدر حالی که داروهایی که گیرنده های هیستامین H2 را مسدود می کنند در درمان زخم معده و اثنی عشر موثر هستند.
ایجاد داروهایی با تأثیر انتخابی ، تا حد زیادی باعث کاهش مصرف بالینی آنها شده است ، زیرا این داروها به میزان قابل توجهی کاهش می یابد اثر جانبی.
اساس مکانیسم عملکرد داروها ، به عنوان یک قاعده ، توانایی آنها در شروع (تحریک) فرآیندهای پیچیده بیوشیمیایی و / یا بیوفیزیکی است که در نهایت باعث تغییر و / یا بهینه سازی فعالیت عملکرد سلول هدف می شوند. داروها می توانند اثر خود را بر روی اندام ها و / یا بهینه کنند. یا سلولهای هدف توسط: تعامل شیمیایی مستقیم؛ تعامل فیزیکی و شیمیایی بر روی غشای سلولی. اقدامات مربوط به آنزیم های تخصصی؛ اقدامات مربوط به ژنهای نظارتی؛ اقدامات روی گیرنده های خاص.
اثر متقابل شیمیایی مستقیم داروها.
این مکانیسم عملکرد داروها بسیار نادر است و می تواند در خارج از سلول ، به عنوان مثال ، در لومن معده یا روده ها تحقق یابد. جوهر آن در این واقعیت نهفته است که داروها با یک مولکول و / یا یونهایی که در بدن در یک وضعیت عادی یا آسیب شناختی در بدن شکل می گیرند وارد یک واکنش شیمیایی مستقیم می شوند. نمونه ای از تعامل شیمیایی مستقیم ، واکنش شیمیایی خنثی سازی اسید هیدروکلریک معده هنگام مصرف داروهای ضد اسید است.
اثر متقابل فیزیکی و شیمیایی داروها بر روی غشای سلولی.
یکی از کارکردهای اصلی غشای سیتوپلاسمی ، اجرای تبادل یونی بین سیتوپلاسم و محیط خارج سلولی است. تبادل یونی transmembrane همچنین می تواند از طریق کانال های یون یونی وابسته به ولتاژ - سدیم ، پتاسیم ، کلسیم ، کلر و غیره انجام شود. برخی از داروها با رسیدن به غشای سلولی با این کانالها ارتباط برقرار می کنند و فعالیت عملکردی آنها را تغییر می دهند. به عنوان مثال ، عمل ضد آریتمیک داروی کلاس IA - کینیدین - بر اساس توانایی آن در مسدود کردن عبور یون های Na + از طریق کانال های سدیم ترانسمیبن است.
تأثیر داروها بر آنزیمهای تخصصی.
مقدار نسبتاً کمی از داروها با تغییر فعالیت برخی از آنزیمهای سلولی تخصصی ، اثر دارویی آن را درک می کنند. به داروهای افزایش فعالیت آنزیمهای سلولی گفته می شود سلفآنزیم ها چنین عملی ، به عنوان مثال ، با قرص های خواب آور و داروی ضد تشنج فنوباربیتال ، که به طور قابل توجهی فعالیت آنزیم های کبدی میکروزومی را تقویت می کند ، وجود دارد. اهمیت بیولوژیکی این تأثیر داروهای فنوباربیتال و داروهای مرتبط در زیر مورد بحث قرار خواهد گرفت.
به داروهای مهار کننده فعالیت آنزیم های تخصصی گفته می شود مهارکننده هاآنزیم ها بنابراین ، به عنوان مثال ، یک داروی ضد افسردگی از گروه مهار کننده های مونوآمین اکسیداز (MAO) ، داروی پیرلیندول با مهار فعالیت آنزیم MAO در سیستم عصبی مرکزی ، اثر ضد افسردگی خود را تحقق می بخشد. توانایی مهار فعالیت آنزیم استیل کولین استراز اساس فعالیت دارویی داروهای آنتی کولین استراز ، به عنوان مثال فیزیوستیگمین است. مشخص شده است که در شرایط فیزیولوژیکی ، استیل کولین استراز غیرفعال می شود (از بین می برد) استیل کولین ، انتقال دهنده عصبی است که باعث تحریک تحریک در سیناپس های سیستم عصبی پاراسمپاتیک می شود. Physostigmine ، با سرکوب فعالیت استیل کولین استراز ، باعث افزایش تجمع در سیناپسهای سیستم پاراسمپاتیک از استیل کولین انتقال دهنده عصبی می شود ، در نتیجه ، میزان صدای سیستم عصبی پاراسمپاتیک افزایش می یابد ، که در سطح سیستمیک بواسطه ایجاد برادی کاردی ، کاهش فشار خون و فشار خون (فشار خون افزایش می یابد) مردمک و غیره داروها می توانند با آنزیم ها برگشت پذیر و برگشت ناپذیر داشته باشند. به عنوان مثال ، داروی انالاپریل به طور معکوس فعالیت آنزیم تبدیل کننده آنژیوتانسین را مهار می کند ، که به ویژه باعث کاهش فشار خون می شود ، در حالی که مواد سمی ارگانوفسفر باعث برگشت ناپذیر فعالیت استیل کولین استراز می شوند. تأثیر داروها بر روی ژنهای نظارتی.در حال حاضر ، دانشمندان در تلاشند تا داروهایی را ایجاد کنند که اثرات دارویی آنها را با تأثیر مستقیم بر فعالیت فیزیولوژیکی ژنهای نظارتی تأثیر بگذارد. به نظر می رسد این روند پس از کشف ساختار ژنوم انسانی در سال 2000 امیدوار کننده است. اعتقاد بر این است که عادی سازی انتخابی عملکرد ژنهای نظارتی تحت تأثیر داروها باعث می شود که در درمان بسیاری از بیماریها از جمله بیماریهایی که قبلاً غیرقابل تحمل بوده اند ، موفقیت حاصل شود. تأثیر داروها بر روی گیرنده ها.قبل از اینکه به ویژگیهای تعامل داروها با گیرنده ها بپردازیم ، لازم است منظور از اصطلاح گیرنده را روشن کنیم (از لاتین). گیرنده - بگیر ، بگیر) از دوره فیزیولوژی مشخص شده است که اصطلاح گیرنده به معنای سازندهای کاملاً تخصصی است که قادر به درک ، تبدیل و انتقال انرژی یک سیگنال خارجی به سیستم عصبی هستند. به این گیرنده ها گفته می شود حسی(از لات. حس - احساس ، احساس ، ادراک). گیرنده های حسی شامل گیرنده های اندام های شنوایی ، بینایی ، بو ، بو ، مزه ، لمس و غیره هستند. گیرنده های حسی این اندام ها متعلق به به اصطلاح بیرون گیرنده ها هستند. اگر از زمان های قدیم وجود ارگان های حسی که به محرک های خارجی تحریک پاسخ می دهند ، وجود گیرنده های حسی در داخل بدن مورد سوال قرار گرفته است. برای اولین بار ، فیزیولوژیست روسی I.F. صهیون ، که در سال 1866 افت فشار خون را به دلیل تحریک آئورت در یک آزمایش خرگوش نشان داد. این کشف باعث جستجو و مطالعه گیرنده های مستقر در بدن شد و خود این گیرنده ها نامیده شدند بینگیرنده ها بهآغاز قرن XX تعداد کافی از گیرنده های حسی آشکار شد و نقش مهم آنها در تنظیم عملکردهای فیزیولوژیکی بدن ثابت شد. در سال 1905 ، J. Langley ثابت کرد که هنگامی که یک دارو بر روی غشای سلولی اعمال می شود ، اگر فقط در یک منطقه خاص از آن استفاده شود ، اثر دارویی ایجاد می شود. علاوه بر این ، این سایت تنها بخش کوچکی از مساحت کل سطح سلول را تشکیل می دهد. این مشاهده به J. Langley اجازه داد که نتیجه گیری کند که سایت های مختلف گیرنده در تعامل با داروها بر روی غشای سلولی وجود دارند. با این حال ، اولویت در ایجاد تئوری گیرنده عمل به داروها متعلق به فیزیولوژیست آلمانی P. P. Erlich است که در سال 1906 برای اولین بار اصطلاح "گیرنده" را معرفی کرد و فرمولاسیون فرضیه را عنوان کرد: "اگر بر روی غشای سلولی ثابت نباشد ، دارو کار نمی کند". طبق تئوری P. Ehlich ، یک مولکول دارویی دارای دو گروه عملکردی فعال است که یکی از آنها تثبیت آن بر روی سطح سلول در ناحیه گیرنده دارو را تضمین می کند ، و گروه دوم عملکردی با گیرنده ارتباط برقرار می کنند و زنجیره پیچیده ای از واکنشهای بیوشیمیایی را ایجاد می کنند که فعالیت فیزیولوژیکی آن (سلول) را تغییر می دهد. . بنابراین ، در اوایل قرن بیستم. آشکار شد که حداقل دو کلاس از گیرنده گیر وجود دارد: گیرنده های حسیانتقال اطلاعات در مورد وضعیت اندام های داخلی و بافت های بدن در سیستم عصبی مرکزی. گیرنده های سلولکه با داروهایی که فعالیت عملکردی سلولهای هدف را تغییر می دهند تعامل دارند.<>بلافاصله لازم به ذکر است که در آینده ، در متن کتاب درسی ، به منظور جلوگیری از سردرگمی در اصطلاحات ، گیرنده های مواد مخدر و مواد بیولوژیکی فعال ، یعنی. سلولی یا cytoreceptoريبا استفاده از اصطلاح "گیرنده" مشخص می شود ، در حالی که گیرنده های حسگر با اصطلاح مشخص کننده فعالیت عملکردی خود ، به عنوان مثال "گیرنده گیر" ، "گیرنده درد" و غیره مشخص می شوند. کشف توسط P. Ehlich در غشای سلولی گیرنده های مواد مخدر به عنوان نقطه شروع برای توسعه علم داروسازی ، به ویژه فارماکودینامیک ، که یکی از اصلی ترین وظایف آن مطالعه مکانیسم های گیرنده عملکرد داروها است. در حال حاضر ، ساختار تعداد زیادی گیرنده سلولی ، ویژگیهای متقابل برخی از ترکیبات بیولوژیکی فعال با آنها آشکار شده است ، که این امر باعث می شود از یک طرف مکانیسم عملکرد داروهای شناخته شده را درک کنیم و از طرف دیگر ، زمینه ساز ایجاد داروهای جدید بسیار مؤثر بود. طبیعتاً ، تصور اینكه در دوره تحول ، گیرنده های داروهای مختلف مصنوعی (شیمیایی به دست آمده) در بدن انسان ایجاد شده است ، به ویژه از آنجایی كه بخش عمده ای از داروهای ارائه شده در بازار دارویی مدرن در 50 سال گذشته یا كمتر سنتز شده اند ، دشوار است. ثابت شده است که دستگاه گیرنده سلول یک سازه کاربردی-ساختاری بسیار باستانی است. بنابراین ، گیرنده های a- و b- آدرنرژیک (گیرنده ها ، اثر متقابل نوراپی نفرین و آدرنالین بر عملکرد عملکردی سلول تأثیر می گذارد) نه تنها در سلولهای حیوانی بلکه در غشاهای سلولی سلولهای گیاهی نیز مشاهده می شود ، به عنوان مثال در سلولهای گیاه نیتلا ، جایی که a- و b- آدرنرژیکها حرکت پروتوپلاسم (محتویات سلولی) را تنظیم می کنند. سپس گیرنده های داروهای کشف شده توسط P. Ehrich چه هستند ، و چرا آنها با آنها ارتباط برقرار می کنند؟ در حال حاضر ، شکی نیست که گیرنده های به اصطلاح دارویی در واقع گیرنده هایی برای مواد فعال بیولوژیکی درون زا (تولید شده در بدن) هستند که در تنظیم فعالیت عملکردی اندام های داخلی و بافت های بدن نقش دارند. چنین ترکیبات فعال بیولوژیکی شامل موادی است که در هنگام انتقال سیگنال عصبی از انتهای عصب رها می شود ، همچنین هورمون ها ، ویتامین ها ، اسیدهای آمینه و غیره. برای هر ماده بیولوژیکی فعال درون زا ، گیرنده های خاصی برای آن وجود دارد. بنابراین ، به عنوان مثال ، ماده بیولوژیکی فعال تولید شده در بدن ، آدرنالین ، می تواند به ویژه خاص را فعال کند - گیرنده های B و آدرنرژیک ، و گلوکوکورتیکواستروئیدها - هورمونهای قشر آدرنال - تنها با گیرنده های گلوکوکورتیکواستروئید ارتباط برقرار می کنند. داروهای مصنوعی که با تعامل با دستگاه گیرنده سلول ، اثرات خود را متوجه می شوند ، در ساختار شیمیایی خود ، کم و بیش شبیه ترکیبات فعال بیولوژیکی درون زا هستند که با گیرنده های مشابه در تعامل هستند. به عنوان مثال ، داروهای فنیل افرین (وازو کانستریکتور مصنوعی) (باعث انقباض عروق) می شوند و فنیل افرین در ساختار شیمیایی خود با ماده فعال درون زا فعال بیولوژیکی نوراپی نفرین نزدیک است ، بنابراین ، مانند نوراپی نفرین ، توانایی تحریک گیرنده های آدرنرژیک را دارد. بعضی اوقات ، به دلیل ویژگی های ساختار شیمیایی آنها ، داروها می توانند نه با خود گیرنده بلکه با قسمت مجاور غشای سلولی ارتباط برقرار کنند. از آنجا که در این حالت ، دارو نه با خود گیرنده بلکه با قسمت مجاور غشای سلولی ارتباط برقرار می کند ، آنها از تأثیر هیجان انگیز یا مسدود کننده بر روی گیرنده صحبت نمی کنند ، بلکه اختصاص دادنثروتمند(از یونانی همه چیز - اثر دیگر یا متفاوت) در نتیجه ممکن است تغییری در ساختار غشای مجاور گیرنده و اجزای فردی گیرنده ایجاد شود که ممکن است منجر به تغییر حساسیت گیرنده به یک ماده بیولوژیکی فعال مخصوص آن شود. در مواردی که حساسیت گیرنده به یک ماده فعال بیولوژیکی افزایش می یابد ، در مورد آن صحبت کنید حساسیت(از لات. حس - احساس) یا در مورد حساسیت(از لات. حساسیت - حساسیت) گیرنده و در مواردی که حساسیت گیرنده کاهش می یابد ، در مورد صحبت کنید حساسیت زداییگیرنده ویژگی خاصیت آلوستریک در این واقعیت نهفته است که داروهایی که دارای این نوع مکانیسم عمل هستند مستقیماً در انتقال یک ضربه عصبی تأثیر نمی گذارند بلکه آن را در جهت مورد نظر اصلاح می کنند. به عنوان مثال ، مکانیسم عمل ضد اضطراب (داروهای ضد اضطراب ؛ مترادف: آرامبخش) ، در ساختار شیمیایی آنها که مشتقات بنزودیازپین است ، مبتنی بر پدیده تحریک آلوستریک گیرنده های بنزودیازپین پس سیناپسی است. تحریک دوم ، به نوبه خود ، باعث تقویت فعال شدن گیرنده های پس سیناپسی مهار کننده اسید گاما آمینوبوتیریک (گیرنده های گابا GABA) می شود که با از بین بردن علائم بیماری های عصبی مانند اضطراب ، اضطراب ، ترس و غیره از نظر بالینی تجویز می شود. گیرنده ها ، در تعامل با آنها ، ماده یا دارویی بیولوژیکی فعال از هر طریق وضعیت عملکردی سلول هدف را تغییر می دهند. خاص علاوه بر گیرنده های خاص ، به اصطلاح غیر اختصاصیبرای گیرنده های مواد مخدر. در ادبیات تخصصی پزشکی ، به این گیرنده ها "محل از بین رفتن" داروها نیز گفته می شود. با تماس با چنین گیرنده ها ، داروها هیچ اثر بیولوژیکی ندارند ، اما خود آنها بیولوژیکی غیرفعال می شوند. نمونه ای از این نوع گیرنده می تواند به عنوان گیرنده هایی که بر روی پروتئین های پلاسما قرار دارند ، بخصوص پروتئین های محلول در آب - آلبومین - خدمت کند. ساختار گیرنده ها کاملاً پیچیده است ، اما بیشتر آنها پروتئین ماکرومولکول ها یا گلیکوپروتئین ها هستند که ممکن است شامل یون ها ، لیپیدها ، اسیدهای نوکلئیک و غیره نیز باشند. گیرنده یعنی. ماکرومولکول پروتئین تشکیل دهنده آن با استفاده از یک ترکیب خاص و خاص برای هر گیرنده ، ترتیب مکانی گروههای شیمیایی آن است. ماکرومولکول پروتئینی تشکیل دهنده گیرنده می تواند در لایه لیپید غشای سیتوپلاسمی یا در داخل سلول بصورت یکپارچه (غوطه ور) شود. کارکرد اصلی گیرنده سلولی این است که یک سیگنال شیمیایی را که از طریق یک ماده فعال بیولوژیکی فعال درون زا و / یا داروها به آن منتقل می شود "بشناسد" و آن را به یک پاسخ سلولی بیوشیمیایی و / یا بیوفیزیکی مربوط تبدیل کند. پیش از این اعتقاد بر این بود كه داروها یا مواد بیولوژیکی فعال درون زا با گیرنده های نوع "كلید و قفل" ارتباط برقرار می كنند ، یعنی. گیرنده دارای ساختاری است که به دارو اجازه می دهد تا گیرنده "شما" را پیدا کند ، به آن متصل شود و مانند گذشته "روشن" شود و "خاموش" شود. با این حال ، با پیشرفت علم پزشکی ، آشکار شد که این کاملاً درست نیست. در حال حاضر ، فرآیندهای مولکولی تبدیل سیگنالهای خارج سلولی به عملکرد سلولی تنظیم کننده داخل سلولی ، قبلاً کاملاً مورد مطالعه قرار گرفته است. مکانیسم هایی که منجر به اثر متقابل مواد درون زا و بیولوژیکی فعال مواد مخدر با گیرنده ها می شوند. هنگام تعامل با گیرنده یک ماده فعال بیولوژیکی درون زا و / یا یک داروی فعال مانند آن اتفاق می افتد ترکیب- تغییر مکانی به شکل ماکرومولکول پروتئین ، که ماشه ای برای فرآیندهای درون سلولی مختلف است که پاسخ یک سلول هدف به یک واسطه و / یا دارو را تعیین می کند. به عنوان مثال ، فعال شدن گیرنده های بتا 2-عضلات صاف برونشها تحت تأثیر فنوترول b2-adrenostimulator باعث افزایش فعالیت آنزیم آدنیلات سیکلاز ، که به تجمع مونوفسفات سیکللیک آدنوزین (cAMP) در سلول و در نتیجه آرام سازی سلول منجر می شود. به طور کلی از نظر بیولوژیکی ، گیرنده های سلولی می توانند به عنوان "ارگان های حسی" کاملاً تخصصی در نظر گرفته شوند که از طریق آنها "اطلاعات" را که مثلاً از سیستم عصبی مرکزی و / یا سیستم غدد درون ریز ناشی می شود ، درک می کنند. با وجود نقش مهم دستگاه گیرنده ، گیرنده ها تنها بخش ناچیزی از غشای سلولی را اشغال می کنند. به عنوان مثال ، دستگاه گیرنده M- کولینرژیک یک سلول بیش از 1/6000 مساحت آن را اشغال نمی کند. بررسی خصوصیات تعامل داروها با گیرنده ، از یک سو به ما این امکان را می دهد تا اساس مکانیسم مولکولی عمل آن را بفهمیم و از طرف دیگر ، اطلاعاتی را در مورد اینکه چه عواملی باید در ساختار داروها ایجاد شود را فراهم می کند تا توانایی آن در تعامل با این گیرنده را تقویت کند ، یعنی. اجازه می دهد تا از ترکیب داروهای جدید بسیار مؤثر استفاده شود. تحت شرایط فیزیولوژیکی ، گیرنده های مختلف سلولی به طور مستقل عمل نمی کنند ، اما در تعامل مداوم با یکدیگر هستند و از این طریق فعالیت خاص سلول را تنظیم می کنند. به عنوان مثال ، فعال شدن گیرنده های B- آدرنرژیک قلبی توسط نوراپی نفرین درون زای باعث بخصوص افزایش تعداد انقباضات قلبی و فعال شدن گیرنده های M- کولینرژیک سلول های قلبی توسط استیل کولین درون زا ، برعکس باعث کاهش تعداد انقباضات قلب می شود. با کشف گیرنده های پیش و پس از سیناپسی ، سهم بسزایی در درک مکانیسم های گیرنده عمل مواد مخدر داشت. سیچرتها(از یونانی خلاصه داستان - اتصال) یک منطقه تماس تخصصی بین سلولهای عصبی یا سایر ساختارهای هیجان انگیز بدن است که انتقال اطلاعات ورودی و حفظ اهمیت اطلاعاتی آن را تضمین می کند. مطالعه ساختار و نقش عملکردی سیناپسها در اواخر قرن 19 آغاز شد. پس از تاریخ شناس اسپانیایی S. Ramon-i Cajal (S. Ramon در Cajal) حضور یک سیستم انتقال تخصصی در سیستم عصبی مرکزی را پیشنهاد کرد. سیناپس در سال 1897 نامگذاری شد ، زمانی که Ch. Sherrington ، فیزیولوژیست انگلیسی ، این اصطلاح را برای اشاره به منطقه تماس بین سلولهای عصبی پیشنهاد کرد. در حال حاضر ، سه نوع سیناپس وجود دارد: 1) سیناپسهای "الکتریکی" که در آن اطلاعات با انتقال یک سیگنال الکتریکی از غشای پیش سیناپسی منتقل می شوند. به این نوع سیناپس گفته می شود efaps(از یونانی افتادگی - تماس تنگ)؛ 2) سیناپسهای "شیمیایی" که در آن اطلاعات از طریق مواد فعال بیولوژیکی ویژه منتقل می شود - انتقال دهنده های عصبی(از یونانی نورون - عصب و لات. واسطه - واسطه؛ مترادف: واسطه)؛ 3) سیناپسهای "مختلط" که در آن اطلاعات به صورت شیمیایی و برقی منتقل می شوند. اثرات دارویی اکثریت قریب به اتفاق داروهایی که بر عملکرد سیناپسها تأثیر می گذارند با تأثیر آنها در مرحله خاصی از انتقال سیگنال در سیناپسهای شیمیایی تحقق می یابد ، یعنی. در خلاصه نوع دوم. به عنوان یک قاعده ، سیناپسهای شیمیایی توسط انتقال دهنده های عصبی طبقه بندی می شوند که تکانه های عصبی را در آنها منتقل می کنند ، به شرح زیر است: سیناپسهایی که استیل کولین به عنوان واسطه عمل می کنند ، کولینرژیک نامیده می شوند. سیناپسهایی که آدرنالین و نوراپی نفرین به عنوان واسطه عمل می کنند ، آدرنرژیک نامیده می شوند. سیناپسهایی که در آنها ATP و آدنوزین به عنوان واسطه عمل می کنند ، پورینرژیک نامیده می شوند. سیناپسهایی که در آنها گاما آمینوبوتیریک اسید به عنوان واسطه عمل می کند ، GABA-ergic و غیره نامیده می شوند. ساختار سیناپس در حال حاضر به خوبی درک شده است. سیناپس شامل یک فرآیند پیش سیناپسی سلول عصبی (انتهای آکسون) و دستگاهی برای دریافت سیگنال است که بر روی غشای یک سلول مؤثر ("اجرایی") قرار دارد.
فارماکودینامیک - اثرات دارویی ، مکانیسم های عمل ، بومی سازی عمل ، انواع عمل داروها.
اثرات دارویی ماده مخدر - تغییر در فعالیت اندام ها ، سیستم های بدن که ماده ایجاد می کند (برای مثال ، افزایش انقباضات قلب ، کاهش فشار خون ، تحریک فعالیت ذهنی ، از بین بردن ترس و تنش و غیره).
هر ماده باعث ایجاد تعدادی از اثرات دارویی مشخصه آن می شود. در هر حالت ، فقط از برخی اثرات دارویی استفاده می شود که به صورت زیر تعریف می شوند اصلی اثرات اثرات دارویی باقی مانده (استفاده نشده ، نامطلوب) نامیده می شود وثیقه.
مکانیسم های عمل داروها - راه هایی که مواد باعث اثرات دارویی می شوند بسیار متنوع است. گزینه های اصلی مکانیسم های اقدام شامل اقدامات زیر است:
- گیرنده های خاص.
- آنزیمها؛
- کانالهای یونی؛
- سیستم های حمل و نقل.
بیشتر داروها روی این عمل می کنند گیرنده های خاص. این گیرنده ها اغلب توسط مولکول های پروتئین فعال فعال هستند ، تعامل با آنها باعث واکنش های بیوشیمیایی می شود که منجر به اثرات دارویی می شوند.
گیرنده های خاصی در ارتباط با غشای سلولی (غشاء) و گیرنده های داخل سلولی (سیتوپلاسمی ، هسته ای).
گیرنده های غشایی (گیرنده های غشای سیتوپلاسمی) به زیر تقسیم می شوند:
- گیرنده ها مستقیماً به کانال های یونی وصل می شوند.
- گیرنده ها به طور مستقیم به آنزیم ها کونژوگه می شوند.
- گیرنده هایی که با پروتئین های G ارتباط برقرار می کنند.
به گیرنده ها مستقیماً به کانال های یونی وصل می شوند، به ویژه ، گیرنده های Ν-کولینرژیک و گیرنده های گابا A را شامل شوند.
هنگامی که گیرنده های NA- کولینرژیک تحریک می شوند (گیرنده های کولینرژیک حساس به نیکوتین) ، کانال های سدیم به طور مستقیم با آنها باز می شوند. تحریک گیرنده های NA- کولینرژیک منجر به کشف کانال های Na + ، ورود یون های Na + به سلول ، دپلاریزاسیون غشای سلولی و یک اثر هیجان انگیز می شود.
گیرنده های GABA A مستقیماً با کانال های کلر همراه هستند. تحریک گیرنده های GABA A منجر به کشف کانال های Cl ، ورود یون های کلرید ، بیش فعالی شدن غشای سلولی و اثر مهاری می شود.
به گیرنده هایی که به طور مستقیم با آنزیم ها مرتبط هستندشامل ، به ویژه ، گیرنده های انسولین به طور مستقیم به تیروزین کیناز کونژوگه می شوند.
گیرنده های پروتئین G - گیرنده های M- کولینرژیک (گیرنده های کولینرژیک حساس به موسکارینی) ، گیرنده های آدرنرژیک ، گیرنده های دوپامین ، گیرنده های مواد افیونی و غیره.
پروتئین های G ، یعنی پروتئین های اتصال دهنده GTP ، در غشای سلولی بومی سازی می شوند و از زیر واحد های α- ، β- و γ تشکیل می شوند. هنگامی که یک دارو با یک گیرنده در تعامل است ، α زیر واحد پروتئین G به GTP (GTP) متصل می شود و در آنزیم ها یا کانال های یونی عمل می کند.
یک گیرنده با چندین پروتئین G تعامل دارد و هر یک از مجتمع های α زیر واحد پروتئین G با GTP در چندین مولکول آنزیم یا در چندین کانال یونی عمل می کند. بنابراین ، مکانیسم تقویت کننده (تقویت کننده) اجرا می شود: وقتی یک گیرنده فعال می شود ، فعالیت بسیاری از مولکول های آنزیم یا بسیاری از کانال های یونی تغییر می کند.
یکی از اولین کسانی که یافت می شود پروتئین های G همراه با β1- گیرنده های قلب بود. هنگامی که عصب سمپاتیک قلب فعال می شود ، β 1-گیرنده گیر هیجان زده می شوند. آدنیلات سیکلاز از طریق پروتئین G فعال می شود. cAMP از ATP تشکیل می شود ، پروتئین کیناز فعال می شود ، تحت این عمل کانال های Ca2 + فسفریله شده و باز می شوند.
افزایش ورودی یون های Ca2 به سلول های گره سینواتریال مرحله چهارم پتانسیل عمل را تسریع می کند ، فرکانس تکانه های تولید شده افزایش می یابد - انقباضات قلب شایع تر می شود.
کشف کانال های Ca2 در فیبرهای میوکارد در حال کار منجر به افزایش غلظت های Ca2 در سیتوپلاسم می شود (ورودی های Ca2 باعث آزادسازی Ca2 از ردیف سارکوپلاسمی می شود). یون های Ca2 به تروپونین C (یک جزء از تروپونین-تروپومیوزین) متصل می شوند. بنابراین ، اثر مهاری تروپونین-تروپومیوزین بر تعامل آکتین و میوزین کاهش می یابد - انقباضات قلب شدت می یابد (شکل 10).
شکل 10- مکانیسم افزایش و تقویت انقباضات قلبی در هنگام تحریک β 1 گیرنده گیرنده. AC - آدنیلات سیکلاز؛ کامپیوتر - پروتئین کیناز؛ CA - گره سینواتریال؛ Ttm - تروپونین-تروپومیوزین.
وقتی عصب پاراسمپاتیک قلب (اعصاب واگ) فعال می شود ، گیرنده های M2-کولینرژیک هیجان زده می شوند و از طریق پروتئین های G ، آدنیلات سیکلاز مهار می شود - انقباضات قلب ضعیف و ضعیف می شوند (عمدتا انقباضات دهلیزی ضعیف می شوند ، زیرا درونی پاراسمپاتیک بطنها نسبتاً ضعیف است).
بنابراین ، پروتئین های G می توانند اثر تحریک کننده و مهاری بر روی آدنیلات سیکلاز داشته باشند. تحریک پروتئین G به عنوان پروتئین G (تحریک) و پروتئین G مهاری (پروتئین G (مهار) تعیین شد) (شکل 11).
شکل 11- مکانیسم تغییر در دفعات و قدرت انقباضات قلب در هنگام تحریک عصب سمپاتیک و پاراسمپاتیک.
سم وبا پروتئین های C را فعال می کند (این منجر به فعال شدن آدنیلات سیکلاز می شود و در وبا توسط ترشح مایع از طریق اپیتلیوم روده بروز می یابد).
سم Pertussis پروتئین G i را فعال می کند.
هنگامی که گیرنده های کولینرژیک M1 ، گیرنده های کلینرژیک M3 ، گیرنده های α1 از طریق پروتئین های G q برانگیخته می شوند ، فسفولیپاز C فعال می شود ، که به شکل گیری اینوزیتول-1،4،5-تری فسفات و diacylglycerol از فسفاتیدیلینوزول-4،5-دیفسفات .
اینوزیتول-1،4،5-تری فسفات بر روی گیرنده های غشای رتیکولوم سارکوپلاسمی حساس به آن عمل می کند و باعث آزاد شدن یون های Ca2 + از شبکه شبکه سارکوپلاسمی می شود (شکل 12). با تحریک گیرنده های عروقی αι رگ های خونی ، این منجر به انقباض عضلات صاف رگ ها و باریک شدن عروق می شود (شکل 13).

شکل 12. تأثیر فسفولیپاز C بر روی سطح سیتوپلاسمی C a2 +.
حساسیت گیرنده ها به آگونیست ها و تعداد گیرنده ها به طور مداوم در حال تغییر است. بنابراین ، پس از تحریک گیرنده های β 1 آدرنرژیک توسط آگونیست β 1-آدنورسپتور ، فسفریلات توسط یک گیرنده کیناز ویژه ، به پروتئین β-arrestin متصل می شوند و در این مجموعه توانایی تعامل با پروتئین های G (حساس سازی گیرنده) را از دست می دهند. مجتمع گیرنده های β1-آدرنرژیک با β- آگستین توسط سلول اندوسیتوز (درونی گیرنده ها) توسط سلول جذب می شود و توسط اندوسوم ها و لیزوزوم ها اسیر می شود. در آندوزومها ، مولکولهای β 1 -restin از گیرنده هایی که مجدداً در غشای سلولی جدا می شوند جدا می شوند. حساسیت گیرنده به آگونیست ها احیا می شود (حساسیت مجدد گیرنده). در لیزوزوم ها ، تخریب مولکول های گیرنده رخ می دهد (پایین تنظیم) (شکل 14).

شکل 13. مکانیسم انقباض عضلات صاف رگهای خونی هنگام تحریک درونی شدن سمپاتیک. FLS - فسفولیپاز C؛ فیفا 2 - فسفاتیدیلینوزیتول -4،5-دی فسفات. اگر 3 - اینوزیتول-1،4،5-تری فسفات ؛ SR - شبکه سارکوپلاسمی؛ KLCM - کیناز زنجیره های سبک میوزین.

شکل 14. حساسیت زدایی و تنظیم پایین گیرنده های β آدرنرژیک.
به گیرنده های داخل سلولی کورتیکواستروئید و گیرنده های هورمون جنسی شامل می شوند. به طور خاص ، گیرنده های گلوکوکورتیکوئید در سیتوپلاسم سلول ها بومی سازی می شوند. پس از ترکیب گلوکوکورتیکوئیدها با گیرنده های سیتوپلاسمی ، مجتمع گیرنده گلوکوکورتیکوئید - هسته به درون هسته نفوذ کرده و در بیان ژن های مختلف تأثیر می گذارد.
توانایی اتصال مواد به گیرنده ها (تمایل مواد به اتصال به گیرنده ها) با اصطلاح " وابستگی" در رابطه با گیرنده های مشابه ، میل به مواد مختلف می تواند متفاوت باشد. برای توصیف وابستگی ، از نشانگر استفاده کنید پK D لگاریتم منفی ثابت تفکیک است ، یعنی غلظت ماده ای که 50٪ گیرنده ها در آن اشغال شده اند.
فعالیت داخلی - توانایی مواد در تحریک گیرنده ها. تعیین شده توسط اثر داروییمرتبط با فعال شدن گیرنده.
در شرایط عادی ، هیچ ارتباط مستقیمی بین وابستگی و فعالیت درونی وجود ندارد. یک ماده می تواند همه گیرنده ها را اشغال کند و باعث ایجاد یک اثر ضعیف شود و برعکس ، یک ماده می تواند 10٪ از گیرنده ها را اشغال کند و باعث ایجاد حداکثر اثر برای این سیستم می شود.
آگونیست ها - موادی با میل و فعالیت داخلی.
آگونیست های کامل تمایل و فعالیت داخلی حداکثر دارند (حتی می توانند بخشی از گیرنده های خاص را در خود جای دهند) حداکثر تأثیر را برای یک سیستم معین داشته باشد.
آگونیست های جزئی (جزئی) داشتن میل به هم و کمتر از حداکثر فعالیت درونی (حتی اگر 100٪ گیرنده های خاص را اشغال کنند) می توانند باعث کمتر از حداکثر اثرات شوند.
آنتاگونیست هاتمایل به هم دارند ، اما فعالیت درونی ندارند و با عملکرد آگونیست های کامل یا جزئی تداخل دارند (آگونیست ها را از ارتباط با گیرنده ها جدا کنید).
اگر با افزایش دوز آگونیست ، عمل آنتاگونیست از بین برود ، چنین تضادی را رقابتی می نامند.
آگونیست های جزئی می توانند متضاد آگونیست های کامل باشند. در صورت عدم وجود آگونیست کامل ، یک آگونیست جزئی گیرنده ها را تحریک می کند و باعث ایجاد یک اثر ضعیف می شود. هنگام تعامل با یک آگونیست کامل ، یک آگونیست جزئی گیرنده ها را اشغال می کند و عملکرد یک آگونیست کامل را مهار می کند. در این حالت ، اثر آگونیست کامل تضعیف می شود.
به عنوان مثال ، پیندولول ، یک آگونیست β آدرنرژیک جزئی β ، در صورت عدم وجود اثرات عصب سمپاتیک بر روی قلب ، باعث تاکی کاردی ضعیف می شود. اما با افزایش لحن عصب سمپاتیک ، پیندولول به عنوان یک مهارکننده واقعی β عمل می کند و باعث برادی کاردی می شود. این در شرایطی است که پیندولول آگونیست جزئی ، اثر نوراپی نفرین واسطه را که یک آگونیست کامل با توجه به گیرنده های β- آدرنرژیک قلب است ، تضعیف می کند.
آگونیست های آنتاگونیست - موادی که در زیرگروه های گیرنده های مشابه متفاوت عمل می کنند: برخی از زیرگروه های گیرنده را تحریک می کنند ، و دیگران را مسدود می کنند. به عنوان مثال ، nalbuphine ضد درد داروی اثرات متفاوتی در زیرگروه های گیرنده مواد افیونی دارد. Nalbuphine گیرنده های κ را تحریک می کند (و بنابراین حساسیت به درد را کاهش می دهد) ، و بلوک های گیرنده μ (و بنابراین از نظر وابستگی به مواد مخدر خطرناک تر است).
نمونه ای از تأثیر مواد روی آنزیم ها ممکن است عملی از عوامل ضد کولین استراز وجود داشته باشد که استیل کولین استراز را مسدود می کند (آنزیمی که استیل کولین را تجزیه می کند) و در نتیجه عملکرد استیل کولین را تقویت و طولانی می کند.
مواد دارویی شناخته شده ای که باعث تحریک یا انسداد می شوند کانالهای یونی غشای سلولی ، یعنی کانالهایی که به طور انتخابی یون های Na + ، K + ، Ca 2+ (سدیم ، پتاسیم ، کانال های کلسیم) و غیره را هدایت می کنند به عنوان مثال:
بی حسی موضعی کانال های Na + را مسدود می کند.
داروهای ضد آریتمی کلاس I (کینیدین ، \u200b\u200bلیدوکائین) کانال های Na + را مسدود می کنند.
مینوکسیدیل کانالهای K + را فعال می کند.
عوامل هیپوگلیسمی از گروه مشتقات سولفونیل اوره کانالهای وابسته به ATP را مسدود می کنند.
کانالهای وراپامیل ، بلوک نیفدیپین Ca2.
نمونه ای از تأثیر مواد روی سیستم های حمل و نقل ممکن است عملی انجام شود:
Reserpine (بلوک های وزیکولی جذب دوپامین و نوراپی نفرین).
گلیکوزیدهای قلبی (مهار سدیم / K + -ATPase).
ضد افسردگیهای سه حلقه ای (بلوک جذب عصبی معکوس نوراپی نفرین و سروتونین).
مسدود کننده های پمپ پروتون (امپرازول و غیره).
مکانیسم های دیگر عمل ممکن است. به عنوان مثال ، مانیتول دیورتیک با افزایش فشار اسمزی در لوله\u200cهای کلیوی ، دیورز را افزایش می دهد. ماده ضد آترواسکلروتیک - کلستیپول - اسیدهای صفراوی را می چسباند (sequestrates) ، از جذب آنها در روده جلوگیری می کند و بنابراین تشکیل اسیدهای صفراوی از کلسترول در کبد فعال می شود و سطح کلسترول در کبدی کاهش می یابد.
مکانیسم های عملکرد داروهای مختلف تا حد مختلفی مورد مطالعه قرار گرفته است. در فرایند مطالعه آنها ، ایده ها در مورد مکانیسم های عمل نه تنها پیچیده تر می شوند بلکه تغییر قابل توجهی نیز می کنند.
مفهوم " محلی سازی عمل"به معنای جایگاه اصلی عمل برخی از داروهای خاص است. به عنوان مثال ، گلیکوزیدهای قلبی عمدتاً بر روی قلب عمل می کنند.
به مفهوم " انواع عمل»مربوط به اقدامات محلی و کلی (تحلیلگر) ، عمل رفلکساقدام اولیه و ثانویه ، اقدام مستقیم و غیرمستقیم.
نمونه ای از یک عمل محلی ممکن است عمل بی حسی موضعی باشد.
بیشتر داروها دارای یک اثر کلی (جاذب) هستند که معمولاً پس از جذب (جذب) یک ماده در خون و توزیع آن در بدن ایجاد می شود.
هر دو با اقدامات موضعی و با تحریک پذیری ، مواد می توانند گیرنده های حساس مختلفی را تحریک کرده و باعث واکنش های رفلکس شوند.
اثر اصلی یک ماده دارویی اثرات آن است که در هر مورد استفاده می شود. همه اثرات دیگر به عنوان مظاهر عوارض جانبی ارزیابی می شوند.
مواد دارویی می توانند بر روی اندامهای خاصی اعمال شوند اقدام مستقیم. علاوه بر این ، اثر داروها می تواند غیرمستقیم باشد. به عنوان مثال ، گلیکوزیدهای قلبی تأثیر مستقیمی بر قلب دارند ، اما با بهبود عملکرد قلب ، این مواد باعث افزایش خون و عملکرد سایر ارگان ها می شوند (اثر غیرمستقیم).