Velika većina lijekovi ima terapeutski efekat promjenom aktivnosti fizioloških sistema ćelija koje se stvaraju u tijelu tokom evolucije. Pod utjecajem ljekovite tvari u organizmu u pravilu ne nastaje nova vrsta ćelijske aktivnosti, mijenja se samo brzina različitih prirodnih procesa. Inhibicija ili ekscitacija fizioloških procesa dovodi do smanjenja ili pojačanja odgovarajućih funkcija tjelesnih tkiva.
Lijekovi mogu djelovati na specifične receptore, enzime, ćelijske membrane ili direktno komunicirati sa staničnim supstancama. Pojedinosti o mehanizmima djelovanja ljekovite tvari studirao na predmetu opšte ili eksperimentalne farmakologije. U nastavku navodimo samo neke primjere glavnih mehanizama djelovanja lijekova.
Dejstvo na specifične receptore. Receptori su makromolekularne strukture koje su selektivno osjetljive na određena hemijska jedinjenja. Interakcija hemikalija s receptorom dovodi do pojave biohemijskih i fizioloških promjena u tijelu, koje se ispoljavaju u određenom kliničkom efektu.
Lijekovi koji direktno pobuđuju ili povećavaju funkcionalnu aktivnost receptora nazivaju se agonisti, a supstance koje inhibiraju djelovanje specifičnih agonista nazivaju se antagonisti. Antagonizam može biti konkurentan i ne konkurentan. U prvom slučaju, tvar lijeka konkurira prirodnom regulatoru (posredniku) za mjesta vezivanja u specifičnim receptorima. Blokada receptora uzrokovana konkurentnim antagonistom može se eliminirati velikim dozama agonista ili prirodnog posrednika.
Različiti receptori su podijeljeni prema osjetljivosti na prirodne posrednike i njihove antagoniste. Na primjer, receptori osjetljivi na acetilholin nazivaju se holinergičkim, a adrenalinski receptori naziva se adrenergički. U pogledu osjetljivosti na muskarin i nikotin, kolinergički receptori se dijele na osjetljive na muskarin (m-holinergički receptori) i nikotinski osjetljivi (n-holinergički receptori). H-holinergički receptori su heterogeni. Utvrđeno je da njihova razlika leži u osjetljivosti na razne supstance. Postoje n-holinergički receptori smješteni u ganglijima autonomnog nervnog sustava i n-holinergički receptori mišića s prugama. Poznate su različite podvrste adrenergičkih receptora, koje su označene grčkim slovima α1, α 2, β1, β2.
Takođe su izolovani H1 i H2 histamin, dopamin, serotonin, opioidni i drugi receptori.
Uticaj na aktivnost enzima. Neki lijekovi povećavaju ili inhibiraju aktivnost specifičnih enzima. Na primjer, fizostigmin i neostigmin smanjuju aktivnost holinesteraze, koja uništava acetilholin i daju efekte karakteristične za pobuđivanje parasimpatičkog nervnog sistema. Inhibitori monoamin oksidaze (iprazid, nialamid), koji sprečavaju uništavanje adrenalina, povećavaju aktivnost simpatičkog nervnog sistema. Fenobarbital i zixorin, povećavajući aktivnost glukuronil-transferaze u jetri, smanjuju nivo bilirubina u krvi.
Fizičko-hemijski učinak na ćelijske membrane. Aktivnost ćelija nervnog i mišićnog sistema ovisi o protoku jona koji određuju transmembranski električni potencijal. Neki lijekovi mijenjaju transport jona.
Dakle, antiaritmički, antikonvulzivni lijekovi, lijekovi za opću anesteziju.
Direktna hemijska interakcija. Lijekovi mogu direktno komunicirati s malim molekulama ili ionima unutar stanica. Na primjer, etilendiaminetetraoctena kiselina (EDTA) snažno veže olovne ione. Princip neposredne hemijske interakcije u osnovi je korištenja mnogih antidota za hemijsko trovanje. Drugi primjer je neutralizacija klorovodične kiseline antacidima.
Farmakodinamika
Proučava se mehanizam delovanja lekova, kao i njihovi biohemijski i fiziološki efekti. Njeni zadaci uključuju opis kemijskih i fizičkih interakcija između lijeka i ciljane ćelije, kao i puni spektar i ozbiljnost njegovih farmakoloških učinaka. Poznavanje farmakodinamičkih obrazaca omogućava vam odabir pravih lijekova. Farmakodinamičke studije pružaju dublje razumijevanje regulacije biokemijskih i fizioloških procesa u tijelu (Katzung B.G., 1998; Lawrence D.R. et al., 2002).
Djelovanje većine lijekova posreduje njihovim vezanjem na makromolekule u tijelu. Promjena funkcionalnog stanja ovih makromolekula, zauzvrat, pokreće lanac biohemijskih i fizioloških reakcija koje se pretvaraju u farmakološki učinak. Makromolekule s kojima kemikalije stupaju u interakciju nazivaju se receptori. Dakle, sve funkcionalno aktivne makromolekule mogu poslužiti kao receptori za lijekove. Iz ove izjave proizlazi nekoliko važnih posljedica. Prvo, uz pomoć lijekova možete promijeniti brzinu bilo kojeg fiziološkog procesa u tijelu. Drugo, lijekovi mijenjaju samo prirodne fiziološke funkcije ćelije, ne dajući joj nova svojstva.
Receptori
Većina receptora su proteini. Riječ je o receptorima hormona, faktorima rasta, posrednicima, proteinima koji sudjeluju u najvažnijim metaboličkim i regulatornim reakcijama (dihidrofolat reduktaza, acetilkolinesteraza), transportnim proteinima (Na +, K + -ATPase), strukturnim proteinima (tubulin). Ćelijske komponente različitog hemijskog karaktera, kao što su nukleinske kiseline, sa kojima antitumorska sredstva međusobno djeluju, mogu također djelovati kao receptori.
Receptori endogenih regulatornih faktora - hormona, posrednika itd., Imaju farmakološki značaj. Ovi receptori služe kao meta mnogih lijekova, obično djeluju selektivno zbog visoke specifičnosti receptora za endogene ligande. Lijekovi koji, nakon vezivanja na receptor, reproduciraju fiziološki učinak endogenog liganda, nazivaju se aganisti ili stimulansi. Lijekovi koji ne uzrokuju taj efekt, ali inhibiraju vezanje endogenih liganda, nazivaju se antagonisti ili blokatori. Supstance čiji je efekat manje izražen od efekta agonista nazivaju se djelomičnim agonistima. Pripravci koji stabiliziraju receptor u neaktiviranoj konformaciji klasificirani su kao inverzni agonisti.
Strukturna i funkcionalna ovisnost
Hemijska struktura lijeka prilično čvrsto određuje njegov afinitet prema receptorima i unutrašnju aktivnost. Neznatna promjena u kemijskoj strukturi može značajno utjecati na farmakološka svojstva.
Na tome se u velikoj mjeri temelji sinteza novih lijekova. Budući da kemijska modifikacija ne mora nužno utjecati na sva farmakološka svojstva jednako, moguće je poboljšati učinkovitost i sigurnost lijeka, povećati njegovu selektivnost i poboljšati farmakokinetičke karakteristike. Na primjer, mnogi antagonisti hormona i posrednika koji se koriste u klinici sintetišu se hemijskim modifikacijama endogenih supstanci.
Bodovi za prijavu lijekova
Budući da učinak lijekova posreduju receptori, točka primjene lijeka određuje se ne samo karakteristikama njegove distribucije, već i lokalizacijom receptora, a farmakološki učinci ovise o funkcionalnom značaju tih receptora. Farmakološki učinci lijekova čiji su receptori zajednički u mnogim organima i tkivima su različiti. Ako ovi receptori obavljaju funkciju koja je vitalna za stanice, nije samo teško koristiti lijek u terapeutske svrhe, već je i nesiguran. Bez obzira na to, takvi lijekovi mogu biti od velike kliničke važnosti. Dakle, srčani glikozidi, koji se široko koriste kod zatajenja srca, mijenjaju transport jona kroz staničnu membranu, o čemu ovisi vitalna aktivnost ćelije. Imaju uzak terapeutski raspon i vrlo su toksični. Drugi primjer su antitumorska sredstva. Ako su receptori s kojima lijek djeluje prisutni na samo nekoliko vrsta diferenciranih ćelija, njegov učinak je selektivniji. Ovi lijekovi mogu imati manje nuspojava, ali ipak, ti \u200b\u200blijekovi mogu biti toksični ako njihovi receptori obavljaju vitalnu funkciju. Neki biološki otrovi (botulinum toksin, itd.) Djeluju na sličan način. Pored toga, čak i ako je izravni farmakološki učinak selektivan, njegove posljedice mogu biti raznovrsnije.
Endogeni receptorski faktorski receptori
Izraz receptor odnosi se na bilo koju makromolekularnu komponentu stanice s kojom se lijek veže. Jedan od najvažnijih receptora za lijekove su ćelijski proteini, koji služe kao receptori za endogene regulatorne faktore - hormone, faktore rasta, posrednike. Vezivanjem na endogeni ligand, receptori prenose signal iz njega u ciljanu ćeliju.
Iz receptora signal stiže do ćelijskih ciljeva (efektorskih proteina) direktno ili posredstvom signalnih molekula - protein-pretvarača. Receptori, proteinski pretvarači i efektorski proteini tvore sistem receptora i efektora. Najbliži efektorski protein u lancu prijenosa signala često nije terminalni efektor (koji direktno utječe na stanične funkcije), već enzim ili transportni protein uključen u stvaranje, transport ili inaktivaciju drugog posrednika - iona ili malog molekula. Drugi posrednik, pak, prenosi informacije na različite unutarćelijske ciljeve, osiguravajući njihov istovremeno reagiranje na signal s jednog receptora.
Receptori, pretvaranje proteina i efektorski proteini ne prenose samo informacije. Oni koordiniraju i signale iz različitih liganda, s jedne strane, i svi ti signali s metaboličkim procesima u stanici, s druge.
Djelujući kao katalizatori, receptori pojačavaju biološki signal. Zbog ovog važnog svojstva služe kao odlične mete za lijekove. Međutim, pojačivači signala nisu samo receptori s enzimskom aktivnošću, već su svi poznati receptori. U stvari, kad se jedan molekul liganda veže na receptor konjugiran na ionski kanal, mnogi ioni prolaze kroz zadnji. Isto vrijedi i za receptore za steroidne hormone: jedna molekula hormona pokreće transkripciju mnogih kopija mRNA, na temelju kojih se sintetišu brojne proteinske molekule.
Ovisno o strukturi i mehanizmu djelovanja, receptore biološki aktivnih tvari dijele se u nekoliko klasa. Broj ovih časova je mali.
Enzimski receptori
Najveća grupa receptora s enzimatskom aktivnošću su membranski receptori sa vlastitom aktivnošću protein kinaze. Fosforiliraju različite efektorske proteine \u200b\u200bkoji se nalaze na unutrašnjoj strani ćelijske membrane. Kao rezultat toga, funkcija ovih proteina ili njihova interakcija sa drugim proteinima se menja.
Postoji još jedna klasa receptora sa aktivnošću proteinske kinaze - to su receptori konjugirani s protein kinazama. Nedostaje im intracelularni katalitički domen, ali prilikom interakcije sa agonistom, vežu se ili aktiviraju unutarćelijske proteinske kinaze na unutrašnjoj površini membrane. Riječ je o receptorima za neurotrofične faktore i receptorima koji prepoznaju antigene za T i B limfocite, a sastoje se od nekoliko podjedinica. Potonji takođe međusobno djeluju sa fosfotirozin fosfatima. Funkcija drugih receptora koji nemaju intracelularnu efektorsku domenu može biti posredovana nekim drugim efektorskim proteinima.
I drugi receptori sa vlastitim enzimatskim djelovanjem imaju sličnu strukturu. Tu spadaju, na primjer, receptori sa vlastitom aktivnošću fosfotirozin fosfataze: njihova izvanstanična domena slična je u slijedu aminokiselina kao adhezijski molekuli. Za mnoge receptore s vlastitom aktivnošću fosfotirozin fosfataze endogeni ligandi nisu poznati. Međutim, prema genetičkim i biohemijskim istraživanjima koja se vrše na različitim tipovima ćelija, enzimska aktivnost ovih receptora igra važnu ulogu. Intraćelijska domena atrijskih receptora natriuretičkih hormona, ostali NUP-ovi, kao i guanilinski receptori, imaju vlastitu aktivnost gvanilat ciklaze i sintetizira cGMP, koji djeluje kao drugi posrednik. Možda postoje i drugi receptori sa vlastitom enzimskom aktivnošću.
Receptori vezani za jonski kanal
Receptori nekih medijatora izravno su povezani s ionskim kanalima, koji djeluju ligandom kako bi selektivno prolazili određene ione kroz staničnu membranu (hemosenzitivni kanali, jonotropni receptorski kanali, ionotropni receptori).
G-proteinski povezani receptori
Ovo je prilično velika klasa receptora koji stupaju u interakciju s efektorima preko G-proteina (proteina koji koriste supstituciju gvanin-difosfata (GDF)) gvanin-trifosfatom (GTP). To uključuje receptore za mnoge biogene amine, molekule lipidnog signala (posebno eikosanoide) i različite peptide bjelančevine i proteini. Enzimi (adenilat ciklaza, fosfolipaza C) i kalijum i kalcijevi membranski kanali djeluju kao efektori. Veliki broj i važna fiziološka uloga receptora spojenih sa G-proteinima čini ih izvrsnim. moji ciljevi za lijekove: otprilike polovina svih lijekova koje su propisali ljekari (osim antibiotika) djeluju na ove receptore.
Stanica može nositi do 20 receptora na svojoj površini od kojih svaki selektivno djeluje s jednom ili više vrsta G-proteina (razlikuju se u različitim vrstama α-podjedinica). Α-podjedinica je u mogućnosti komunicirati s jednim ili više efektorskih proteina, što vam omogućava koordiniranje signala iz receptora različitih liganda koristeći jedan G-protein. S druge strane, jedan receptor može pokrenuti nekoliko mehanizama prenosa unutarćelijskog signala, aktivirajući nekoliko vrsta G-proteina i djelovati na različite efektorske proteine \u200b\u200bkroz istu α-podjedinicu. Tako složen sistem divergencije i konvergencije signala omogućuje fleksibilnu regulaciju staničnih funkcija (Ross, 1992).
Intraćelijski receptori
Receptori steroidnih i štitnih hormona, kalcitriol i retinoidi su topljivi unutarćelijski proteini koji se vežu za DNK koji reguliraju transkripciju određenih gena (Mangelsdorf et al., 1994). Ti receptori pripadaju super-porodici regulatora transkripcije osjetljivih na ligande. Funkcija faktora transkripcije reguliše se fosforilacijom, interakcijom sa ćelijskim proteinima, metabolitima i drugim regulatornim komponentama stanice.
Drugi posrednički sistemi
cAMP. Sekundarni posrednički sustavi također su uključeni u integraciju vanjskih signala. Iako ima mnogo više poznatih receptora i proteinskih signalnih molekula od drugih medijatora, potonji su uključeni u mnoge putove unutar ćelijskog prenosa signala. Najviše proučavani drugi posrednici uključuju cAMP, cGMP, Ca 2+, IF3 (inozitol trifosfat), DAG (diacilglicerol), NO. Ova grupa heterogenih spojeva konstantno raste. Drugi posrednici stupaju u interakciju direktno (mijenjajući međusobni metabolizam) ili indirektno (djelujući na iste unutarćelijske ciljeve). Funkcija drugog medijatora, kao i regulacija njihovog formiranja (ili oslobađanja), cijepanja i izlučivanja iz stanice, prikladno se razmatra na primjeru cAMP. Ovaj drugi medijator sintetizira se pod utjecajem adenilat citaze aktiviranjem mnogih receptora konjugiranih s G-proteinima. G s protein aktivira adenylat ciklazu, G i protein inhibira.
Postoji najmanje 10 izoformnih adenilata ciklotaza, specifičnih za tkivo, koji se razlikuju u mehanizmima regulacije aktivnosti.
U pravilu, cAMP aktivira protein kinaze A (cAMP-ovisne proteinske kinaze), malu grupu srodnih proteina. Ove proteinske kinaze, zauzvrat, fosforiliraju ne samo konačne unutarćelijske mete (enzimi, transportni proteini), već i druge proteinske kinaze i druge regulatorne proteine. Potonje uključuje, na primjer, faktore transkripcije. Oni su odgovorni za cAMP-posredovanu regulaciju transkripcije gena, pružajući odgođeni stanični odgovor na signal. Uz aktiviranje protein kinaza, cAMP djeluje izravno na kanale kationskih membrana, koji imaju važnu ulogu, posebno, u funkcioniranju neurona. Stoga signal iz cAMP-a uzrokuje lanac biohemijskih promjena u ciljanoj ćeliji.
Kalcijum Drugi dobro proučeni drugi posrednik je unutarćelijski Ca 2+. Ioni Ca 2+ ulaze u citoplazmu na različite načine: duž membranskih kanala (ovisno o G-proteinima, o naponu ovisnih, reguliranih K + ili Ca-Ca 2+), kao i putem kanala smještenih u posebnim područjima endoplazmatskog retikuluma i otvaranja pod djelovanjem AKO 3 i u skeletnim mišićima kao rezultat depolarizacije membrane. Uklanjanje kalcijuma iz citosolne plazme događa se na dva načina: apsorbuje ga endoplazmatski retikulum ili se izlučuje iz ćelije. Ca 2+ odašilje signale mnogo većem broju proteina nego cAMP - enzimi koji su uključeni u stanični metabolizam, protein kinaze, proteini koji vezuju kalcijum. Potonji interaktivno djeluju s ostalim završnim i intermedijarnim učincima.
Regulacija receptora
Receptori ne samo da kontroliraju fiziološke i biohemijske funkcije, već i služe kao objekti regulacije. Ova regulacija provodi se na nivou sinteze i razgradnje njihovih makromolekula, stvaranjem kovalentnih veza s drugim molekulama, interakcijom sa regulatornim proteinima i kretanjem receptora. Pretvorba proteina i efektorskih proteina je takođe podložna regulaciji. Regulatorni signali mogu poticati iz intracelularnih putova prenosa aktiviranih stimulacijom samog receptora (preko povratnog mehanizma), kao i iz drugih receptora (direktno ili indirektno).
Dugotrajna stimulacija receptora za lijekove obično dovodi do smanjenja reakcije na njega - pri istoj koncentraciji lijek izaziva manje izražen efekat. Ovaj fenomen, nazvan desenzibilizacija, vatrostalni, zarazni, igra važnu ulogu u kliničkoj praksi: na primjer, s dugotrajna upotreba β-adrenergički agonisti za liječenje pacijenata sa AD ozbiljnost reakcije na ove lijekove je smanjena.
Homološka desenzibilizacija odnosi se samo na stimulirane receptore i specifična je za ligand. S heterološkom desenzibilizacijom, smanjuje se ozbiljnost reakcije na druge ligande, čiji receptori djeluju istim putem prenosa signala unutar staničnog signala. U prvom slučaju negativne povratne informacije daju se učinkom na sam receptor (fosforilacija, proteoliza, smanjena sinteza), u drugom slučaju osim receptora može utjecati i na ostale proteine \u200b\u200bkoji su uključeni u prijenos unutarćelijskog signala.
Naprotiv, ako se receptori dugo ne stimulišu, njihova osjetljivost na agoniste se povećava (na primjer, kod dugotrajnog liječenja β-adrenoblokatorom propronololom povećava se osjetljivost β-adrenergičkih receptora na β-adrenostimulanse).
Poremećaji zbog oslabljene funkcije receptora
Pored individualnih razlika u osjetljivosti na lijekove, postoje i bolesti uzrokovane disfunkcijom određenih komponenti mehanizma prijenosa unutarćelijskog signala s receptora na efektor. S gubitkom funkcije visoko specijaliziranih receptora, fenotipske manifestacije bolesti mogu biti ograničene (na primjer, feminizacija testisa povezana s genetskom odsutnosti ili strukturnim oštećenjima androgenih receptora). Ako se krši univerzalniji mehanizam unutar prijenosa staničnog signala, simptomi bolesti su raznolikiji, kao što su, na primjer, miastenija gravis i neki oblici dijabetesa melitusa otpornog na inzulin, uzrokovani autoimunim disfunkcijama N-holinergičkih receptora i inzulinskih receptora. Defekti u bilo kojoj komponenti koja je uključena u prijenos signala s mnogih receptora dovode do više endokrinih poremećaja. Primjer je heterozigotni oblik nedostatka proteina G s koji aktivira adenilat ciklazu u svim stanicama (Spiegel i Weinstein, 1995). Homozigotni oblik nedostatka ovog proteina vjerovatno će rezultirati smrću.
Poremećaji u strukturi ili lokalizaciji receptora mogu se pokazati kao oslabljena ili pojačana reakcija na lijek, kao i drugi neželjeni efekti.
Mutacije koje kodiraju genske receptore mogu mijenjati i odgovor na jednokratnu upotrebu lijeka i efikasnost dugotrajnog liječenja. Na primjer, nedostatak β-adrenergičkih receptora odgovornih za opuštanje glatkih mišića bronha i reguliranje otpornosti dišnih puteva pogoršava smanjenje osjetljivosti ovih receptora na β-adrenostimulanse tijekom dugotrajnog liječenja pacijenata s AD. Kad se utvrde mutacije odgovorne za oslabljenu funkciju receptora i kloniraju odgovarajući geni, bit će moguće razviti metode liječenja takvih bolesti.
Klasifikacija receptora
Tradicionalno su receptori za lijekove identificirani i klasificirani na temelju učinaka i relativne aktivnosti selektivnih agonista (stimulansa) i antagonista (blokatora) koji djeluju na te receptore. Na primjer, efekti acetilkolina koji se reproduciraju pri interakciji s holinergičkim receptorima muskarin alkaloida i blokirani su atropinom nazivaju se muskarinima, a učinci koji se reproduciraju u interakciji s holinergičkim receptorima nikotina nazivaju se nikotinskim učincima. Receptori koji posreduju učinke muskarina i nikotina nazivaju se M i N holinergičkim receptorima. Iako takva klasifikacija obično ne odražava mehanizam djelovanja lijekova, prikladno je za sistematizaciju njihovih učinaka. Zapravo, tvrdnja da lijek stimulira receptore određene vrste, istodobno određuje spektar djelovanja ovog lijeka i tvari koje te efekte pojačavaju ili slabe. Međutim, valjanost takvih tvrdnji može se promijeniti prepoznavanjem novih tipova i podvrsta receptora, otkrivanjem dodatnih mehanizama djelovanja lijekova ili prethodno nepoznatih nuspojava.
Podtipovi receptora
Sa pojavom sve većeg broja vrlo selektivnih lijekova postalo je jasno da se prethodno poznate vrste receptora dijele na brojne podvrste. Metode molekularnog kloniranja postale su značajna pomoć u proučavanju novih podtipova receptora, a priprema rekombinantnih receptora olakšala je stvaranje lijekova koji selektivno djeluju na te receptore. Različite ali povezane podtipove receptora često (iako ne uvijek) komuniciraju s različitim agonistima i antagonistima. Receptori za koje nisu identificirani selektivni agonisti ili antagonisti, obično ne pripadaju jednoj podtipi, već izoformama istog receptora. Odvojeni podtipovi mogu se razlikovati u mehanizmima prijenosa unutarćelijskog signala. M1 i M3 holinergički receptori, na primjer, djeluju putem proteina q q, koji aktivira fosfolipazu C, neizravno uzrokujući oslobađanje Ca2+ iz unutarćelijskih depoa, i M2 i M4 holinergičke receptore preko proteina G i koji inhibira adilatnu ciklazu. Istodobno, podjela receptora na vrste i podvrste često se određuje ne mehanizmom djelovanja, već slučajnim odabirom ili se temelji na ustaljenim idejama. Dakle, α 1 -, α 2 - i β-adrenergički receptori se razlikuju u odgovoru na lijekove i u prenosu signala (aktiviraju proteine \u200b\u200bG i, G q i G s, respektivno), iako su α i β-adrenergički receptori različitih vrsta, i α1 - i α2-adrenoreceptori - za različite podvrste unutar iste vrste. Izoforme α 1 -adrenoreceptora α 1A, α 1B i α 1D malo se razlikuju u svojim biohemijskim svojstvima; isto je karakteristično za podtipove izoformraze β-adrenergičkih receptora (β 1, β 2 i β 3).
Razlike između receptorskih podtipova koriste se za stvaranje visoko selektivnih lijekova, na primjer, lijekovi koji imaju različito djelovanje na isto tkivo zbog vezanja na receptore podtipa koji se razlikuju u mehanizmima prijenosa unutarćelijskih signala. Osim toga, lijekovi mogu selektivno ciljati određene stanice ili tkiva koja eksprimiraju receptore podtipa. Što je veća selektivnost lijekova (u odnosu na određeno tkivo ili u odnosu na određeni učinak), povoljniji je omjer njegovih koristi i neželjenih efekata.
Korištenjem molekularno-genetskih metoda otkriveni su ne samo različiti izoformi receptora, već i geni koji kodiraju nove, do tada nepoznate receptore. Mnogi od tih receptora su već dodijeljeni jednoj ili drugoj poznatoj klasi, a njihova funkcija je proučavana pomoću odgovarajućih liganda. Međutim, ligandi još uvijek nisu pronađeni za neke receptore.
Otkrivanje mnogih izoforma istog receptora kodiranih različitim genima (posebno ako se izoformi ne razlikuju u mehanizmima intracelularnog prijenosa signala i interakciju s istim endogenim ligandima) omogućava da se ekspresija receptora u različitim stanicama neovisno regulira u skladu s potrebama tijela u različitim starosnim periodima.
Djelovanje lijekova bez receptora
Nisu svi lijekovi djeluju kroz makromolekularne strukture - receptore. Neki lijekovi djeluju u interakciji s malim molekulima ili ionima koji su normalno prisutni u tijelu ili u jednom ili drugom patološkom stanju. Dakle, antacidi neutraliziraju solnu kiselinu u stomaku. Mesna (lijek koji se bubrezima brzo izlučuje i neutralizira slobodne radikale) veže se na aktivne metabolite nekih lijekova protiv raka, smanjujući ozbiljnost neželjenih reakcija iz mokraćovoda. Broj biološki neaktivnih tvari (na primjer manitol) može se unijeti u količinama dovoljnim da se poveća osmolarnost bioloških tekućina i na taj način promijeni raspodjela izvanstanične i unutarćelijske tekućine. Uz pomoć ovih tvari moguće je povećati diurezu, povećati bcc, ukloniti moždani edem. Pored toga, upotrebljavaju se kao laksativi.
Neki lijekovi se mogu integrirati u komponente ćelije i mijenjati njihove funkcije zbog strukturnih sličnosti s tvarima koje čine ove komponente. Na primjer, analozi purina i pirimidina ubačeni su u nukleinske kiseline i koriste se kao antivirusna i antitumorska sredstva.
A.P. Viktorov "Klinička farmakologija"
U pravilu se mehanizam djelovanja lijekova temelji na njihovoj sposobnosti pokretanja (pokretanja) složenih biokemijskih n / ili biofizičkih procesa koji u konačnici mijenjaju i / ili optimiziraju funkcionalnu aktivnost ciljne ćelije.
Lijekovi mogu provesti svoje djelovanje protiv organa i / ili ciljnih stanica putem:
Izravna hemijska interakcija;
Fizičko-hemijska interakcija na staničnoj membrani;
Djelovanje na specijalizirane enzime;
Akcije na regulatorne gene;
Djelovanje na specifične receptore.
Direktna hemijska interakcija LS. Taj mehanizam djelovanja lijekova je prilično rijedak i može se realizirati izvan ćelije, na primjer, u lumenu želuca ili crijeva. Njegova suština leži u činjenici da lijekovi ulaze u direktnu kemijsku reakciju s molekulima i / ili ionima koji se u tijelu formiraju u normalnom stanju kada dođe do patološkog stanja. Primjer izravne kemijske interakcije je kemijska reakcija neutralizacije klorovodične kiseline želuca pri uzimanju antacidnih lijekova (vidjeti T. 2, str. 112).
Fizičko-hemijska interakcija lijekova na staničnu membranu. Jedna od glavnih funkcija citoplazmatske membrane je provođenje ionske razmjene između citoplazme i izvanćelijskog okruženja. Transmembranska ionska razmjena može se odvijati i kroz posebne transmembranske ionske kanale koji ovise o naponu - natrijum, kalijum, kalcijum, hlor itd. Neki lijekovi, dospijevajući do ćelijske membrane, stupaju u interakciju sa tim kanalima i mijenjaju njihovu funkcionalnu aktivnost. Tako se, na primjer, antiaritmički učinak lijeka klase IA, kinidin, temelji na njegovoj sposobnosti da blokira prolazak Na + jona kroz transmembranske natrijumske kanale (vidi T. 2, str. 35).
Učinak lijekova na specijalizovane enzime. Relativno mala količina lijekova ostvaruje njegov farmakološki učinak mijenjanjem aktivnosti nekih specijaliziranih ćelijskih enzima. Lijekovi koji povećavaju aktivnost staničnih enzima nazivaju se induktorima enzima. Takvim djelovanjem raspolažu, na primjer, tablete za spavanje i antikonvulzivni lijek fenobarbital, koji značajno pojačava aktivnost mikrosomalnih enzima jetre. Biološki značaj ovog efekta fenobarbitala i njemu bliskog LS bit će razmatran u nastavku.
Lijekovi koji inhibiraju djelovanje specijaliziranih enzima nazivaju se inhibitorima enzima. Tako, na primjer, antidepresiv iz skupine inhibitora monoamin oksidaze (MAO), lijek pirindol ostvaruje svoj antidepresivni učinak inhibiranjem aktivnosti enzima MAO u središnjem živčanom sustavu (vidjeti T. 1, str. 294).
Sposobnost inhibiranja aktivnosti enzima acetilkolinesteraza osnova je farmakološkog djelovanja antiholinesteraznih lijekova, na primjer fizostigmina. Poznato je da u fiziološkim uslovima acetilholinesteraza inaktivira (uništava) acetilholin, neurotransmiter koji prenosi ekscitaciju u sinapsama parasimpatičkog nervnog sistema. Fizostigmin, suzbijajući aktivnost acetilholinesteraze, pospješuje nagomilavanje u sinapsama parasimpatičkog sistema neurotransmitera acetilkolina, uslijed čega se povećava ton parasimpatičkog živčanog sustava, što se očituje na sistemskom nivou razvojem bradikardije, snižavanja krvnog tlaka (zglobnog tlaka, lučenja krvi (mokraćnog sustava), lučenja (krvožilnog trakta) (zglobni tlak) (mokraćni trakt), zglobni tlak (mokraćni trakt), zglobni tlak (mokraćni sustav), lučenje krvnih stanica (lučenje krvnih stanica) učenik itd.
Lijekovi mogu djelovati reverzibilno i nepovratno s enzimima. Na primjer, lijek enalapril reverzibilno inhibira aktivnost enzima koji pretvara angiotenzin, što posebno povlači za sobom smanjenje krvnog pritiska, dok toksične supstance organofosfor nepovratno inhibiraju aktivnost acetilkolinesteraze.
Učinak lijekova na regulatorne gene. Trenutno naučnici pokušavaju stvoriti lijekove koji ostvaruju svoje farmakološke učinke direktno utječući na fiziološku aktivnost regulatornih gena. Ovaj se trend čini posebno obećavajući nakon što je 2000. godine dešifrirana struktura ljudskog genoma. Vjeruje se da će selektivna normalizacija funkcije regulatornih gena pod utjecajem lijekova omogućiti postizanje uspjeha u liječenju mnogih, uključujući i ranije neizlječivih, bolesti.
Učinak lekova na receptore. Prije nego što pređemo na specifičnosti interakcije lijekova s \u200b\u200breceptorima, potrebno je razjasniti što podrazumijevamo pod pojmom „receptor“ (od latinskog recipio - uzmi, uzmi).
Iz tijeka fiziologije poznato je da pojam "receptor" znači visoko specijalizirane formacije koje su u stanju opažati, transformirati i prenositi energiju vanjskog signala u živčani sustav. Takvi se receptori nazivaju senzornim (od lat. Sensus - osjećaj, osjet, percepcija).
Senzorni receptori uključuju receptore organa sluha, vida, mirisa, ukusa, dodira itd. Senzorni receptori ovih organa pripadaju takozvanim eksteroreceptorima.
Ako je prisutnost osjetilnih organa koji reagiraju na vanjske podražaje iritacije poznata još od davnina, prisustvo senzornih receptora unutar tijela dovodilo se u pitanje do sredine 19. stoljeća. Prvi put je prisustvo takvih receptora u organizmu sugerirao ruski fiziolog I.F.Pion, koji je 1866. godine pokazao pad krvnog tlaka zbog iritacije aorte u pokusu sa kunićima. Ovo je otkriće dovelo do pretraživanja i proučavanja receptora smještenih unutar tijela, a ti su se sami receptori nazivali interoreceptori.
Početkom 20. vijeka otkriven je dovoljan broj senzornih interoreceptora i dokazana njihova važna uloga u regulaciji fizioloških funkcija tijela.
Godine 1905. J. Langley je dokazao da se, kada se lijek nanosi na staničnu membranu, razvija farmakološki učinak ako se primjenjuje samo na određeno područje nje. Pored toga, ovo mesto čini samo mali deo ukupne površine ćelijske površine. Ovo opažanje je omogućilo J. Langleyju da zaključi da na staničnoj membrani postoje specijalizirana mjesta receptora koja su u interakciji s lijekovima.
Međutim, prioritet u stvaranju teorije receptora za djelovanje lijekova pripada njemačkom fiziologu P. Ehrlichu, koji je 1906. prvi put uveo termin "receptor" i formulirao postulat "lijek ne djeluje ako nije fiksiran na staničnoj membrani". Prema teoriji P. Ehrlicha, molekula lijeka ima dvije funkcionalno aktivne skupine, od kojih jedna osigurava njegovo fiksaciju na staničnoj površini u regiji receptora za lijek, a druga funkcionalna skupina djeluje u interakciji s receptorom i pokreće složen lanac biohemijskih reakcija koje mijenjaju njegovu (staničnu) fiziološku aktivnost .
Dakle, već početkom 20. stoljeća. postalo je očigledno da postoje najmanje dvije klase interoreceptora: senzorni receptori koji prenose informacije o stanju unutrašnjih organa i tjelesnih tkiva u središnji živčani sustav; označavanje receptora koji stupaju u interakciju sa lijekovima koji mijenjaju funkcionalnu aktivnost ciljnih stanica.
Odmah treba napomenuti da bi se u budućnosti u tekstu udžbenika, kako bi se izbjegla konfuzija u terminologiji, receptori za lijekove i biološki aktivne tvari, tj. označeni ili citoreceptori. označit će se terminom "receptor", dok će senzorni interoreceptori biti označeni terminom koji karakterizira njihovu funkcionalnu aktivnost, na primjer, "baroreceptori", "receptori bola" itd.
Otkriće P. Ehrlicha na staničnoj membrani receptora za lijekove poslužilo je kao polazna osnova za razvoj farmakološke znanosti, posebice farmakodinamike, čiji je jedan od glavnih zadataka proučavanje receptorskih mehanizama djelovanja lijekova.
Trenutno su otkrivene struktura velikog broja staničnih receptora, značajke interakcije određenih biološki aktivnih spojeva s njima, što je omogućilo, s jedne strane, razumijevanje mehanizma djelovanja poznatih lijekova, a s druge strane, to je bila osnova stvaranja novih vrlo učinkovitih lijekova.
Naravno, teško je zamisliti da su se tijekom evolucije u ljudskom tijelu formirali receptori za različite sintetičke (hemijski dobivene) lijekove, posebno jer je velika većina lijekova predstavljenih na modernom farmaceutskom tržištu sintetizirana u posljednjih 50 godina ili manje. Dokazano je da je receptorski aparat ćelije vrlo drevna funkcionalno-strukturna formacija. Dakle, a- i β-adrenoreceptori (receptori, čija interakcija norepinefrin i adrenalin utječu na funkcionalnu aktivnost ćelije) nalaze se ne samo u životinjskim ćelijama, već i na staničnoj membrani biljnih stanica, na primjer, u stanicama biljne nittele, gdje su a- i β- adrenorecentori regulišu kretanje protoplazme (ćelijski sadržaj).
Koji su onda receptori za lijekove koje je otkrio P. Ehrlich i zašto s njima interakciju?
Trenutno nema sumnje da su takozvani receptori lijekova zapravo receptori za endogene (proizvedene u tijelu) biološki aktivne tvari uključene u regulaciju funkcionalne aktivnosti unutarnjih organa i tjelesnih tkiva. Takvi biološki aktivni spojevi uključuju tvari koje se oslobađaju iz živčanih završetaka u vrijeme prijenosa živčanog signala, kao i hormone, vitamine, aminokiseline itd. Za svaku endogenu biološki aktivnu tvar postoje strogo specifični receptori za nju. Tako, na primjer, biološki aktivna supstanca proizvedena u tijelu, adrenalin, može aktivirati strogo specifične a- i β-adrenoreceptore, a glukokortikosteroidi - hormoni nadbubrežne kore - u interakciji su samo s glukokortikosteroidnim receptorima koji su im strogo specifični.
Sintetski lijekovi koji svoj učinak ostvaruju interakcijom s receptorskim aparatom stanice, po svojoj kemijskoj strukturi manje su više slični endogenim biološki aktivnim spojevima koji stupaju u interakciju sa sličnim receptorima. Na primjer, sintetički vazokonstrikcijski lijekovi (koji uzrokuju vazokonstrikciju) lijekovi fenilefrin su po svojoj kemijskoj strukturi bliski endogenoj biološki aktivnoj tvari norepinefrinu, stoga, poput norepinefrina, ima sposobnost stimulacije a-adrenoreceptora.
Ponekad, zbog osobitosti njihove kemijske strukture, lijekovi mogu komunicirati ne sa samim receptorom, već sa susjednim dijelom stanične membrane. Budući da u ovom slučaju, lijek djeluje ne sa samim receptorom, već sa susjednim dijelom stanične membrane, oni ne govore o uzbudljivom ili blokirajućem efektu na receptor, već o alosteričnom (od grčkog alios - još jedan, drugačiji) učinak, odnosno učinku. Kao rezultat toga, može doći do promjene u strukturi membrane pored receptora i pojedinačnih komponenti samog receptora, što može podrazumijevati promjenu osjetljivosti receptora na biološki aktivnu tvar specifičnu za njega. U slučajevima kada se povećava osetljivost receptora na biološki aktivnu supstancu, oni govore o senzibilizaciji (od latinskog sensus - smisao) ili osetljivosti (od latinskog sensibilis - osetljivost) receptora, a u slučajevima kada se osetljivost receptora smanjuje, oni govore o desenzibilizaciji receptor.
Posebnost alosteričnog efekta leži u činjenici da lijekovi koji imaju ovakav mehanizam djelovanja ne utječu izravno na prijenos živčanog impulsa, već ga modificiraju u željenom smjeru. Na primjer, mehanizam djelovanja anksiolitika (lijekovi protiv anksioznosti; sinonim: sredstva za smirenje), koji su po svojoj kemijskoj strukturi derivati \u200b\u200bbenzodiazepina, zasnovan je na fenomenu alosteričnog pobuđivanja postinaptičkih receptora benzodiazepina. Pobuđenje potonjih, pak, potiče aktiviranje inhibicijskih postsinaptičkih receptora gama-aminobuterne kiseline (GABA receptora), što se klinički očituje eliminacijom simptoma neurotičnih bolesti kao što su anksioznost, anksioznost, strah, itd.
Receptori, u interakciji s kojima biološki aktivna tvar ili lijek na bilo koji način mijenja funkcionalno stanje ciljne ćelije, nazivaju se specifičnim.
Pored specifičnih receptora, izolirani su takozvani receptori specifični za lijek. U specijaliziranoj medicinskoj literaturi ovi receptori se nazivaju i "mjesto gubitka" lijekova. Kontaktom s takvim receptorima lijekovi nemaju nikakvog biološkog učinka, ali sami postaju biološki neaktivni. Primjer ove vrste receptora mogu poslužiti kao receptori smješteni na proteinima plazme, posebno na proteinima topivim u vodi - albuminu. Značaj ovog fenomena bit će detaljno razmotren u nastavku (vidi T. 1, str. 72).
Struktura receptora je prilično složena, ali većina je proteinskih makromolekula ili glikoproteina koji mogu uključivati \u200b\u200bione, lipide, nukleinske kiseline itd. Receptor tj. proteinu makromolekulu koja ga tvori odlikuje se specifičnim, specifičnim za svaki receptor, prostornim rasporedom njegovih hemijskih grupa. Proteinska makromolekula koja tvori receptor može se integrirati (uroniti) u lipidni dvosloj citoplazmatske membrane ili lokalizirati unutar ćelije. Glavna funkcija ćelijskog receptora je da „prepozna“ hemijski signal koji mu se prenosi putem endogene biološki aktivne supstance i / ili lijekova i transformiše ga u odgovarajući biohemijski i / ili biofizički odgovor ćelije.
Ranije se vjerovalo da lijekovi ili endogene biološki aktivne tvari djeluju u interakciji s receptorima tipa "ključ i bravica", tj. receptor ima takvu strukturu koja omogućava lijeku da nađe "vaš" receptor, poveže se s njim i, po njemu, "uključi" i "isključi". Međutim, razvojem medicinske nauke postalo je očito da ego i nije baš tako. Trenutno su molekularni procesi pretvorbe izvanstaničnih signala u unutarćelijsku, regulirajući staničnu funkciju, već prilično proučeni. mehanizmi koji rezultiraju djelovanjem interakcije endogenih biološki aktivnih tvari ili lijekova s \u200b\u200breceptorima.
Pri interakciji s receptorom endogene biološki aktivne tvari i / ili aktivnog L C poput njega, dolazi do konformacije - prostorne promjene u obliku proteinske makromolekule, koja je pokretač različitih procesa unutar ćelijskih stanica koji određuju odgovor ciljne stanice na posrednika i / ili lijeka. Na primjer, aktiviranje bronhijalnih adrenergičkih receptora glatkih mišića pod utjecajem β 2 -adrenostimulatora fenoterola dovodi do povećanja aktivnosti enzima adenilat ciklaze, što doprinosi nakupljanju cikličkog adenosinofonofata (cAMP) u stanici i, kao rezultat,, staničnom opuštanju.
U opštem biološkom smislu, ćelijski receptori se mogu smatrati strogo specijalizovanim „čulnim organima“ ćelija, kroz koji oni opažaju „informacije“ koje potiču, na primer, iz centralnog nervnog sistema i / ili endokrinog sistema. Uprkos važnoj ulozi receptora, receptori zauzimaju samo neznatan dio stanične membrane. Na primjer, M-holinergički receptorski aparat stanice zauzima ne više od 1/6 000 svoje površinske površine.
Proučavanje karakteristika interakcije lijekova s \u200b\u200breceptorom, s jedne strane, omogućava nam razumijevanje osnova molekularnog mehanizma njegovog djelovanja, a s druge strane, daje podatke o tome koje bi promjene trebale biti učinjene u strukturi lijekova kako bi se povećala njegova sposobnost interakcije s ovim receptorom, tj. . omogućava ciljanu sintezu novih vrlo efikasnih lijekova.
U fiziološkim uvjetima različiti ćelijski receptori ne funkcioniraju neovisno, ali su u stalnoj interakciji jedni s drugima, čime reguliraju specifičnu aktivnost stanice. Na primjer, aktiviranje srčanih β-adrenergičkih receptora endogenim norepinefrinom uzrokuje, posebno, porast broja srčanih kontrakcija i aktiviranje M-holinergičkih receptora srčanih stanica endogenim acetilkolinom, naprotiv, uzrokuje smanjenje broja srčanih kontrakcija.
Otkrivanjem pre- i postsinaptičkih receptora velik je doprinos razumijevanju mehanizama djelovanja lijekova. Sinapsija (iz grčkog sinapsis - veza, komunikacija) je specijalizirana kontaktna zona između živčanih ćelija ili drugih uzbudljivih struktura tijela koja osigurava prijenos dolaznih informacija i očuvanje njenog informacijskog značenja. Proučavanje strukture i funkcionalne uloge sinapsi započelo je krajem 19. vijeka. nakon toga španski histolog S. Ramon n Cajal (S. Ramon kod Cajala) predložio je prisustvo specijalizovanog prenosnog sistema u centralnom nervnom sistemu. Sinapsi su svoje ime dobili 1897. godine, kada je engleski fiziolog C. Sherrington predložio ovaj termin da se odnosi na kontaktno područje između nervnih ćelija.
Trenutno postoje tri vrste sinapsi:
1) "električne" sinapse u kojima se informacije prenose prenošenjem električnog signala sa pre-sinaptičke membrane. Ova vrsta sinapse naziva se efaps (od grč. Ephapsis - tijesan kontakt);
2) "hemijske" sinapse u kojima se informacije prenose putem posebnih biološki aktivnih tvari - neurotransmitera (od grč. Neuron - nerv i lat. Mediator - medijator; sinonim: medijator);
3) „mješovite“ sinapse u kojima se informacije prenose i hemijski i električno.
Farmakološki učinci ogromne većine lijekova koji utječu na funkcije sinapsi ostvaruju se njihovim djelovanjem na goti ili drugi stupanj prijenosa signala u hemijskim sinapsama, tj. u sinapsama druge vrste.
Po pravilu, hemijske sinapse se klasifikuju od neurotransmitera koji u njima prenose nervne impulse:
Sinapse u kojima acetilkolin djeluje kao posrednik nazivaju se holinergičkim;
Sinapse u kojima adrenalin i norepinefrin djeluju kao posrednik nazivaju se adrenergički;
Sinapse u kojima ATP i adenozin djeluju kao posrednik nazivaju se purinergički;
Sinapse u kojima gama-aminobuterna kiselina djeluje kao posrednik nazivaju se GABA-ergičke itd.
Struktura sinapse je trenutno dobro razumljiva. Sinapsa se sastoji od presinaptičkog procesa nervne ćelije (akson-kraj) i aparata za prijem „signala“ koji se nalazi na membrani efektorske („izvršne“) ćelije.
Akson eferentnog neurona, približavajući se efektorskoj ćeliji, gubi mijelinsku ovojnicu, širi se i formira takozvano presinaptičko zadebljanje (Sl. 1.5). Površina živaca koji završava prema staničnoj membrani efektorske ćelije naziva se presinaptička membrana. Mjesto ćelije efektora nasuprot presinaptičkoj membrani naziva se postsinaptička membrana (vidi Sliku 1.5). Ovisno o strukturnim značajkama sinapse, presinaptička membrana može imati više ili manje nabora i na taj način ima veće ili manje područje. U hemijskim sinapsama, presinaptička membrana ne dolazi u direktni kontakt s postsinaptičkom membranom, već je odvojena od nje malom udaljenošću koja se naziva sinaptički rascjep (vidi Sl. 1.5).
Presinaptičko zadebljanje, tj. krajnji dio aksona sadrži veći broj mitohondrija, unutarćelijskih organela koje sudjeluju u sintezi i akumulaciji energije što je veće od tijela neurona, što ukazuje na intenzitet energetskih procesa koji se događaju u ovom dijelu živčane stanice. Osim mitohondrija, presinaptička zadebljanja sadrže veliki broj malih vezikula - vezikula. U prosjeku se nalazi oko 20.000 vezikula u jednom prezinaptičkom zadebljanju. Potonji su smješteni neravnomjerno u presinaptičkom zadebljanju, u pravilu se većina nalazi u blizini presinaptičke membrane. Neurotransmiter se sintetiše u tijelu i aksonu neurona i akumulira se u vezikulama. Svaka vezikula sadrži nekoliko hiljada molekula neurotransmitera (od I 000 do 50 000). Kada se dogodi nervni impuls, vezikula se stapa s presinaptičkom membranom i neurotransmiter se izlučuje u sinaptičku pukotinu (vidi Sliku 1.5).
Sl. 1.5. Shematski dijagram strukture "hemijske" sinapse:
a je shematska slika; b - elektronska mikrografija; 1- presinaptički nervni završetak; 2 - prssinaptička membrana; 3 - postsinaptička membrana; 4 - sinaptička pukotina; B - vezikula; NM - neurotransmiter; P - postsinaptički receptor: OZ - "obrnuti" hvatanje neurotransmitera; SF je specijalizovani enzim koji uništava višak neurotransmitera u sinaptičkoj pukotini
Funkcionalno aktivne tvorbe receptora smještene su na postsinaptičkoj membrani koja je u stanju interakcije s neurotransmiterima koji se oslobađaju iz presinaptičke membrane tokom prolaska živčanog impulsa. Receptori smješteni na postsinaptičkoj membrani nazivaju se sinaptičkim ili postsinaptičkim receptorima u specijaliziranoj medicinskoj literaturi. Pod postsinaptičkim receptorima podrazumijevaju se makromolekule proteinske prirode ugrađene u postsinaptičku membranu s genetski unaprijed određenom strukturom i funkcijom, sposobne za reverzibilnu interakciju s neurotransmiterima i / ili lijekovima zbog funkcionalnih skupina aktivnog centra („prepoznavajući“ dio makromolekule).
Prijenos živčanog signala u sinapsi je sljedeći: pod utjecajem živčanog podražaja vezikule se kreću prema presinaptičkoj membrani, a neurotransmiter se egzocitozom izlučuje u sinaptičku cijev (vidi Sl. 1.5). Neurotransmiter koji se oslobađa u sinaptičku pukotinu dopire do postsinaptičke membrane, gdje, u interakciji s postsinaptičkim receptorom, pokreće lanac biokemijskih i / ili biofizičkih reakcija, čiji je rezultat fiziološki odgovor ciljne ćelije. Međutim, ne sva količina oslobođenog neurotransmitera dostiže postsinaptičke receptore i s njima komunicira. Deo neurotransmitera zarobljava presinaptička membrana i „vraća se“ na mesta skladištenja. Ovaj fenomen se naziva fenomen ponovnog preuzimanja neurotransmitera.
Preostala količina neurotransmiterskog receptora koji ne reagira na interakciju uništava se u sinaptičkoj pukotini specijalizovanim enzimima. Taj fenomen se naziva degradacija neurotransmitera. Na primjer, enzim acetilkolinesteraza katalizira (ubrzava) proces razgradnje (uništavanja) u sinaptičkoj pukotini neurotransmitera acetilkolina.
Za razliku od neurotransmitera, njegovi metabolički proizvodi imaju aktivnost neurotransmitera. Čitav proces interakcije neurotransmitera sa receptorima i uništavanje njegovog viška specifičnim enzimom izuzetno je kratak i ne prelazi 2 ms (1 ms \u003d 0,001 s).
Tako kratko trajanje ovog procesa objašnjava se, s jedne strane, izuzetno brzim oslobađanjem neurotransmitera iz receptora, a s druge, velikom brzinom enzimske inaktivacije neurotransmitera u sinaptičkoj skloništu.
Temeljno funkcionalna aktivnost sinapse može se promijeniti na sljedeći način:
Da biste ubrzali, smanjili ili blokirali sintezu, akumulaciju i / ili katabolizam (uništavanje) neurotransmitera u presinaptičkom završetku. Kao rezultat toga, sadržaj neurotransmitera i, kao posljedica toga, intenzitet njegove fiziološke aktivnosti nekako će se promijeniti.
Na primjer, simpatolitički rezerpin sprečava nakupljanje kateholamina u sinaptičkim vezikulama do njihovog potpunog pražnjenja. Kao rezultat toga, količina neurotransmitera norepinefrina oslobođenog u sinaptičku pukotinu naglo pada. Na nivou sistema, ovaj se efekat ostvaruje u obliku pada krvnog pritiska. Neki lijekovi ne utječu izravno na sadržaj neurotransmitera u presinaptičkom završetku, ali inhibiraju aktivnost enzima koji ih uništavaju. Tako djeluje niz antidepresiva. Na primjer, antidepresiv pirlindol inhibira (suzbija) aktivnost enzima monoamin oksidaza u presinaptičkom prestanku i, kao rezultat, povećava koncentraciju neurotransmitera kao što su norepinefrin, dopamin i serotonin u njemu. Klinički se taj učinak pirlindola očituje smanjenjem osjećaja anksioznosti i straha, poboljšanim raspoloženjem, povećanom fizičkom aktivnošću itd .;
Promijenite (olakšajte, komplicirajte) sposobnost neurotransmitera da prodre kroz presinaptičku membranu i, prema tome, povećanjem ili smanjenjem količine neurotransmitera oslobođenog u sinaptičku pukotinu sa svakim pulsom.
Na primjer, psihostimulans amfetamin olakšava oslobađanje kateholamina u adrenergičkim sinapsama središnjeg živčanog sustava i na taj način povećava njihov sadržaj u sinaptičkoj pukotini. Klinički se ovaj učinak lijeka očituje u poboljšanom raspoloženju, osjećaju navale snage, povećanju performansi. Tetanus toksin blokira oslobađanje inhibicijskih neurotransmitera (GABA, glicin) u središnjem živčanom sustavu i na taj način naglo smanjuje njihov sadržaj u sinaptičkoj pukotini, što se klinički očituje razvojem napadaja;
Blokira ili stimulira ponovni unos neurotransmitera od strane presinaptičke membrane i, na taj način, povećava ili smanjuje koncentraciju neurotransmitera u sinaptičkoj pukotini.
Primjerice, triciklički antidepresiv imipramin blokira ponovni unos neurotransmitera norepinefrina presinaptičkom membranom i na taj način naglo povećava njegovu koncentraciju u sinaptičkoj pukotini. Klinički se taj efekat imipramina manifestuje poboljšanim raspoloženjem, povećanom mentalnom i fizičkom aktivnošću;
Stimuliraju ili blokiraju aktivnost enzima koji uništavaju neurotransmiter u sinaptičkoj pukotini.
Na primjer, antiholinesterazni lijek fitostigmin smanjuje aktivnost enzima acetilkolinesteraza, koji uništava neurotransmiter acetilkolin u sinaptičkoj pukotini, a time pridonosi povećanju njegove koncentracije, što se može klinički manifestirati, naročito padom intraokularnog pritiska i suženjem zjenice.
Stimuliraju ili blokiraju postsinaptičke receptore, tj. imitiraju ili blokiraju učinak neurotransmitera.
Na primjer, opojni analgetici koji pobuđuju postsinaptičke opioidne receptore i tako oponašaju utjecaj neurotransmitera - enkefalina. Strihin blokirajući receptore inhibicijskog neurotransmitera glicina ometa realizaciju njegovog inhibicijskog dejstva, kao rezultat toga, strahnin u visokim dozama izaziva napadaje.

Sl. 1.6. Shematski prikaz lokalizacije pre- i postsinaptičkih receptora kao što je prikazano adrenergičkom sinapsom (objašnjenje u
NM - neurotransmiter; M2 (-) - holinergički „inhibicijski“ presinaptički heteroreceptor; β 1 (+) - adrenergički „aktivirajući“ presinaptički autoreceptor; β - adrenergički postsinaptički receptor
Pored receptora koji se nalaze na postsinaptičkoj membrani, tj. postsinaptički receptori, receptori smješteni na presinaptičkoj membrani, tj. presinatičkih receptora (Sl. 1.6). Uprkos činjenici da i pre- i postinaptički receptori mogu pobuđivati \u200b\u200bisti neurotransmiter, funkcionalna uloga ovih receptorskih formacija u sinapsama je različita. Ako su postsinaptički receptori konačna veza za prijenos živčanog impulsa do organa efektora, tj. omogućavaju jednosmerno sprovođenje nervnog impulsa od centra prema periferiji, a zatim preinaptički receptori učestvuju u
regulacija aktivnosti neurotransmitera sinapse, tj. donekle utiču na procese oslobađanja i / ili sinteze neurotransmitera u njemu. Mora se naglasiti da presinaptički receptori ne direktno sudjeluju u provođenju živčanog impulsa od neurona do organa efektora.
Presinaptički receptori dijele se u dvije velike grupe: auto- i heteroneuromodulatorni receptori (vidi Sl. 1.6).
Presinaptički autoreceptori uključuju receptore koje pobuđuje vlastiti neurotransmiter za ovu sinapsu.
Primjerice, u sinapsama lokaliziranim u području kontakta somatskih živaca i prugastih mišića, kad u sinaptičkoj pukotini postoji višak acetilholina, interakcija s presinaptičkim autoreceptorima inhibira oslobađanje novog dijela neurotransmitera. ekscitacija presinaptičkih autoreceptora reguliše otpuštanje acetilkolina iz presinaptičkih terminala.
Međutim, na presinaptičkoj membrani, pored autoreceptora, tj. receptora koji su osjetljivi na neurotransmiter koji prenose ekscitaciju u određenoj sinapsi, mogu se nalaziti receptori koji nisu osjetljivi na neurotransmiter koji odašilju ekscitaciju u određenoj sinapsi, ali koji djeluju na drugi tip neurotransmitera.
Na primjer, na sinaptičkoj presinaptičkoj membrani, u kojoj je acetilholin neurotransmiter, mogu se nalaziti presinaptički receptori osjetljivi na neurotransmiter norepinefrin. Ova vrsta presinaptičkog receptora naziva se heteroneuromodulirajući receptor.
Dakle, sinapsa je složena anatomska i funkcionalna formacija koja osigurava prijenos nervnog impulsa s neurona na neuron ili iz neurona u efektorsku ćeliju.
Slijed funkcionalne aktivnosti sinapse (faze sinaptičkog prijenosa) je sljedeći:
Sinteza i akumulacija neurotransmitera u vezikulama lokaliziranih u presinaptičkim zadebljanjima (sinteza neurotransmitera se događa ne samo u presinaptičkim zadebljanjima, već i u neuronima i aksonima);
Oslobađanje neurotransmitera u sinaptičku pukotinu u trenutku prolaska nervnog impulsa;
Interakcija neurotransmitera sa postsinaptičkim receptorima, što povlači za sobom aktiviranje receptora i promjenu funkcionalne aktivnosti efektorske ćelije;
Neaktivacija neurotransmitera (enzimatsko) i / ili njegovo povratanje presinaptičkom membranom, tj. obnavljanje sposobnosti sinapse da ponovo prenosi živčani impuls u efektorsku ćeliju.
Sinapsi imaju sljedeća osnovna svojstva:
Jednostrano provođenje pobude (živčani impuls može proći samo iz presinaptičke membrane u postsinaptičku);
Sinaptičko kašnjenje, tj. određeno vrijeme se troši na prijenos nervnog impulsa u sinapsi. (Brzina sinaptičke transmisije u prosjeku je više od 10 puta manja od brzine širenja nervnog impulsa kroz živac. Za hemijsku sinapsu obično se kreće od 0,2 do 0,5 ms);
Umor - postepeno smanjenje ili potpuni prestanak prenosa nervnog impulsa s dugotrajnom nervnom stimulacijom. Osnova ovog fenomena je, s jedne strane, iscrpljivanje rezervi neurotransmitera u presinaptičkim zadebljanjima, a s druge strane, pad osjetljivosti na postsinaptičke receptore na neurotransmiter;
Velika osjetljivost sinaptičkih formacija na lijekove i otrove.
Na posljednjem svojstvu sinapsi temelji se cjelokupna farmakologija lijekova koji utječu na funkcionalnu aktivnost sinapsi smještenih u različitim organima i tkivima tijela. Mora se naglasiti da objekt farmakološkog djelovanja može biti bilo koji od faza sinaptičkog prijenosa. Kao lijekovi koji utječu na sinaptički prijenos koriste se egzogeni analozi neurotransmitera, njihovih kemijskih prekursora i druge biološki aktivne tvari koje na bilo koji način mogu promijeniti funkcijsku aktivnost sinapse.
Treba napomenuti da mnogi lijekovi imaju ne jednu već nekoliko točaka primjene djelovanja na razini sinapse. Tako, na primjer, antidepresiv pirlindol ne samo da inhibira aktivnost enzima monoamin oksidaze u sinaptičkoj pukotini, već i blokira ponovni unos norepinefrina presinaptičkom membranom.
U odnosu na lokalizaciju receptora u sinapsi, oni se mogu podijeliti na presinaptičke, postsinaptičke i ekstrasynaptičke. Potonji, na primer, uključuju receptore koji se nalaze na staničnim membranama trombocita.
Sa stanovišta ćelijske topografije (lokacije), receptori se također mogu klasificirati prema njihovoj lokaciji na staničnim strukturama, kako slijedi:
membranski receptori - receptori smješteni na citoplazmatskoj membrani;
citosolni receptori - receptori smješteni na unutarćelijskim formacijama;
nuklearni receptori - receptori smješteni na membrani ćelijskog jezgra.
Kao što je ranije napomenuto, kao rezultat interakcije s receptorom endogenih bioloških supstanci ili lijekova, mijenja se funkcionalna aktivnost ciljnih stanica. Taj se postupak može provesti na različite načine, strogo definirani za različite vrste receptora. Sukladno s tim, trenutno se razlikuju četiri vrste receptora od kojih svaki ima svoj, u osnovi različit od ostalih, mehanizam kojim signal od receptora pokreće kaskadu biokemijskih i / ili biofizičkih reakcija koje vode promjeni funkcionalnog stanja ciljnih stanica.
Prve tri vrste receptora lokalizirane su na staničnoj (citoplazmatskoj) membrani, a četvrta vrsta receptora uključuje citosolne i nuklearne receptore.
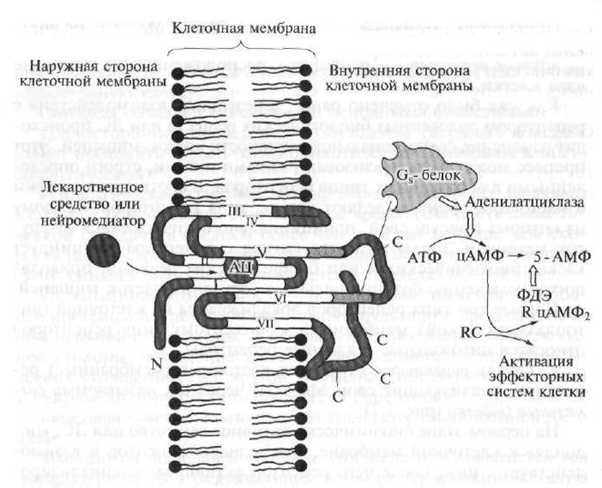
Receptori tipa I uključuju stanične (membranske) receptore koji svoje efekte ostvaruju kroz takozvane signalne G-proteine \u200b\u200b(Sl. 1.7).
U prvom stupnju biološki aktivna tvar ili lijek, koji se "uspinje" do stanične membrane, "prepoznaje" receptor i reagira s njim, nakon čega receptor aktivira specijalizirani signalni G protein koji se nalazi na unutarnjoj površini membrane. Nadalje, aktivirani G-protein mijenja funkcionalnu aktivnost unutarnjeg efektorskog elementa, koji su u pravilu enzimi. Tada efektorski element, koji je enzim, aktivira sekundarni glasnik, ili sekundarni glasnik, što pokreće kaskadu biohemijskih reakcija koje mijenjaju funkcionalnu aktivnost ciljnih stanica.
Stanični receptori tipa I, tj. receptori spojeni sa signalnim G-proteinima su strukturno slični jedni drugima, a po svojoj prostornoj organizaciji oni su serpentinska (od francuskog serpantina - zmija, lopta) strukture (Sl. 1.8).
Sl. 1.7. Struktura receptora tipa 1 (objašnjenje u tekstu)
Sl. 1.8. Shematski prikaz strukture "serpentina"
receptor:
N je polipeptidni dio receptora smješten iznad ćelijske membrane; C je polipeptidni dio receptora smješten ispod ćelijske membrane; AC - aktivno središte receptora s kojim lijek djeluje; ide neurotransmiter; ATP - adenozin trifosfat - sekundarni glasnik; cAMP - ciklični adenozin monofosfat; 5-AMP - adenozin-5 "monofosfat; PDE - fosfodiesteraza; R. RC - enzim zavisan od cAMP (protein kinaza) sa regulatornim i katalitičkim (ubrzavanjem reakcije) podjedinice; 1-VII - polipeptidni lanci serpentinskog receptora
Serpentinski receptori uključuju naborane polipeptidne lance (polipeptid je spoj visoke molekulske težine koji je lanac međusobno povezanih aminokiselinskih ostataka) koji sedam puta prodiru u staničnu membranu.
Endogene biološki aktivne tvari ili lijekovi mogu se vezati za takozvani "džep" formiran polipeptidnim lancem i smješten u debljini stanične membrane, što povlači za sobom aktiviranje signala, koji se prenosi u dijelove lanca receptora smještene u citoplazmi ćelije. Signal G proteini u interakciji sa cy-
stolni (unutarćelijski) dijelovi poda i peptidni lanac; aktivirati i pokrenuti kaskadu biokemijskih reakcija koje mijenjaju njegovu funkcionalnu aktivnost u odnosu na ciljanu ćeliju, tj. pokrenuti primarni farmakološki odgovor.
Trenutno je poznato nekoliko vrsta signalnih G proteina.
Signal G, -proteini. Ti signalni proteini u pravilu aktiviraju efektorski element - enzim adenilat ciklazu, koji zauzvrat potiče sintezu u ćeliji (iz ATP-a) sekundarnog glasnika - cikličnog adenosinofenofosfata (cAMP). Biološka uloga cAMP-a kao sekundarnog glasnika je veoma važna. Na primjer, povećanje njegovog sadržaja u srčanim ćelijama povlači za sobom povećanje učestalosti i snage srčanih kontrakcija. Uz to, povećanje koncentracije cAMP u različitim ciljnim ćelijama uzrokuje opuštanje glatkih mišića krvnih žila i bronha, mobilizaciju energetskih rezervi (propadanje ugljikohidrata u jetri), suzbija sposobnost agregacije trombocita, snižava tonus miometrija (mišića materice) i mokraćnog mjehura itd.
Brojni neurotransmiteri, poput adrenalina (aktiviranjem β-adrenergičkih receptora), dopamina (aktiviranjem D1-dopaminskih receptora), adenozina (aktiviranjem adenosinskih A2 receptora) pripadaju endogenim biološki aktivnim tvarima sa sposobnošću aktiviranja proteinskih signala Gs. histamin (aktiviranjem histamina G2 -receptorop), serotonin (aktiviranjem serotoninskih 5-HT 4-receptora), kao i niza hormona, na primjer, vazopresin (stimulacijom V2-vasopresin receptora) itd.
Signal G i -proteini. Za razliku od signalnih G s proteina, aktivacija signalnih proteina G i ne stimulira, ali inhibira, aktivnost efektorskog elementa, enzima adenilata ciklaze, što dovodi do smanjenja koncentracije cAMP u ciljnim ćelijama sekundarnog glasnika. Smanjenje sadržaja cAMP-a u ciljanim ćelijama uzrokuje smanjenje srčanih kontrakcija, povećanje tonusa krvnih žila i bronha, tj. suprotan učinak na povećanje sadržaja cAMP-a na ciljane ćelije. Pored toga, određeni broj signalnih G i proteina uključen je u regulaciju funkcionalne aktivnosti transmembranskih jonskih Ca2+ i K + kanala.
Brojni neurotransmiteri, na primjer, adrenalin i norepinefrin (aktiviranjem 2-adrenoreceptora), dopamin (aktiviranjem D2 - dopaminskih receptora), adenozin (aktiviranjem A1, pripadaju endogenim biološki aktivnim tvarima sa sposobnošću aktiviranja signalnih G i-proteina. adenozinski receptori), acetilkolin (aktiviranjem M2 i M4 muskarinskih receptora) itd.
Signal G ^ proteini. Ti signalni proteini doprinose aktivaciji drugog efektorskog elementa ciljane ćelije, enzima fosforilaza C, koji zauzvrat potiče stvaranje sekundarnih glasnika dijacilglicerola (DAG) i inozitola 1,4,5-trifosfata (ITP) u ciljanim ćelijama. Prvi od njih (DAG) povezan je s staničnom membranom i pokreće biohemijske reakcije uključene u regulaciju kontraktilnog statusa, rast i dijeljenje stanica i izlučivanje određenih hormona ciljanim stanicama. Pod utjecajem enzima fosfolipaza A2, DAG se može metabolizirati u arahidonsku kiselinu, koja sudjeluje u sintezi biološki aktivnih tvari poput eikosanoida - prostaglandini, prostaciklini, tromboksani, leukotriene (vidjeti T. I, str. 478).
Drugi sekundarni glasnik, ITF, nije fiksiran na staničnoj membrani i prelazi se u unutarćelijski medij (citosol), gdje pokreće oslobađanje Ca 2+ iona iz staničnih depoa, tj. promoviše prelazak neaktivnih Ca 2+ jona u aktivni oblik.
Mnogi istraživači smatraju ione Ca 2+ tercijarnim glasnikom, ili posrednikom. To je zbog činjenice da je uloga iona Ca 2+ u regulaciji funkcionalne aktivnosti ćelija vrlo važna. Ioni Ca 2+ mogu ući u ćeliju iz vanjskog okruženja kroz posebne transmembranske ionske kanale i / ili se oslobađati iz depoa ćelije. Glavni depo (mesto akumulacije neaktivnih Ca 2+ iona) u ćeliji je endoplazmatski, ili sarkoplazmatski retikulum (retikulum sarkoplazmatični unr, sinonim: endoplazmatski retikulum - unutarćelijski organela, koji je sistem tubula i cisterni smještenih u citoplazmi; ograničena je dijelom koji pruža citoplazmu, koja je ograničena dijelom u citoplazmi; transport supstanci u citoplazmi). Slobodni (aktivni) Ca 2+ ioni koji dolaze iz sarkoplazmatskog retikuluma u citoplazmu uzajamno djeluju s nekim Ca2+ vezujućim proteinima, od kojih je najvažniji kalmodulin. Kompleksni "kalmodulin-Ca 2+" i / ili kompleksi Ca2+ iona sa drugim proteinima koji vezuju kalcijum pokreću kaskadu biohemijskih reakcija u ćeliji. Kao rezultat, ovisno o ciljanim organima u kojima se događa taj proces, započinje povećanje kontraktilne funkcije miokarda i skeletnog mišića, povećanje tonusa glatkih mišića krvnih žila, bronha i maternice, povećanje sekretorne aktivnosti žljezdanog tkiva, stimulacija oslobađanja neurotransmitera iz živčanih završetaka itd. . Dokazano je i da Ca 2+ ioni imaju sposobnost povećanja aktivnosti enzima koji su uključeni u metabolizam proteina, ugljikohidrata i masti.
Pored izravne veze između sekundarnih glasnika - DAG i ITF i, stoga, signalnih G q proteina, ioni Ca 2+ u fiziološkim uvjetima imaju prilično složenu interakciju s cAMP sekundarnim glasnikom, čija aktivnost regulira signal G i G i proteini. Tako je pokazano da slobodni ioni Ca 2+ koji ulaze u citoplazmu živčane ćelije putem sistema kalmodulin-Ca 2+ pokreću smanjenje sadržaja iAMP u ćeliji. Istovremeno, za održavanje otvorenog stanja kalcijevih jonskih kanala u ćeliji potrebne su visoke koncentracije cAMP-a, tj. smanjenje sadržaja cAMP inicirano kompleksom kalmodulin-Ca 2+ povlači za sobom prestanak unosa slobodnih Ca2+ jona u citoplazmi. S druge strane, postoje dokazi da cAMP sekundarni glasnik povećava apsorpciju slobodnih Ca 2+ jona sarkoplazmatskim retikulumom, tj. potiče prelazak Ca 2+ iona iz slobodnog, aktivnog oblika u vezani, neaktivni oblik.
Kao rezultat povećanja sadržaja sekundarnih glasnika u ciljnim ćelijama - DAG i ITF - povećava se tonus glatkih mišića, povećava se sekrecija žlijezda, olakšava se oslobađanje neurotransmitera iz presinaptičkih završetaka, sposobnost agregacije trombocita itd.
Endogene biološki aktivne supstance sa sposobnošću aktiviranja proteina C q signala uključuju neurotransmitere poput norepinefrina (aktiviranjem 1 -adrenoreceptora), acetilkolina (aktiviranjem muskarinskih receptora M1 i M3), serotonina (usled aktivacije serotonina 5-HT2a receptore), histamin (zbog aktiviranja histaminskih H1 receptora), kao i druge endogene biološki aktivne tvari, na primjer, bradikinin i angiotenzin.
Trenutno su pored nabrojanih signalnih G-proteina (G s, G |, G q) identifikovani i drugi signalni G-proteini - G s, G i, G q, čija fiziološka uloga još uvek nije jasna. Ali istovremeno postoje dokazi da je, na primjer, signal C o protein uključen u regulaciju funkcionalne aktivnosti transmembranskih jonskih kanala.
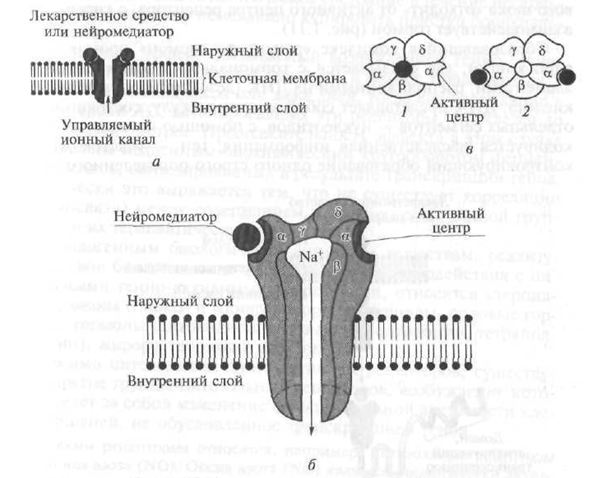
Funkcionalna jedinica receptora tipa 11 je transmembranski (prodire kroz čitavu debljinu stanične membrane) protein (enzim). Sam receptor sastoji se od dva identična fragmenta, koji se zovu monomeri. Monomeri su smješteni na beznačajnoj udaljenosti jedan od drugog, a sam monomer se sastoji od dvije funkcionalno aktivne podjedinice - domene, međusobno povezanih polipeptidnim segmentom koji presijeca membranu lipidnog dvosloja (slika 1.9). A-podjedinica monomera strši iznad vanjske površine membrane i odgovorna je za vezivanje receptora na biološki aktivne tvari, a P-podjedinica je uronjena u citoplazmu stanice.

Sl. 1.9. Struktura receptora tipa II (objašnjenje u tekstu): 1 - a-podjedinica monomera; 2 - β-podjedinica monomera
Nakon vezanja biološki aktivne tvari na α-podjedinicu receptora, receptor se mijenja iz neaktivnog monomernog stanja u aktivno dimerno stanje u kojem se dva monomera kombiniraju u ravnini membrane (vidi sliku 1.9). U tom se slučaju stimulira enzimska aktivnost citoplazmatske β-podjedinice receptora, što rezultira kaskadom biokemijskih reakcija koje mijenjaju njegovo funkcionalno stanje u ciljanoj ćeliji.
Kao transmembranski enzim koji tvori receptor u pravilu se koriste takvi enzimi kao tirozin kinaza ili gvanilat ciklaza.
Primjer receptora za tirozin kinazu su receptori za inzulin (vidjeti T. 1, str. 435).
Put prijenosa signala gvanilat ciklaze započinje interakcijom α-podjedinice receptora s endogenom biološki aktivnom supstancom, na primjer, atrijskim natriuretičkim faktorom (ANF), koji je biološki aktivna tvar koju luče atrijske ćelije i koja sudjeluje u regulaciji srčanih kontrakcija. Kao rezultat ove interakcije dolazi do promjene u konfiguraciji receptora, koja se sastoji u kombiniranju njegovih monomera u dimer. Taj postupak aktivira enzimski receptor koji se nalazi u njegovoj citosolnoj β-podjedinici, tj. enzim gvanilat ciklaza, koji zauzvrat potiče porast koncentracije cikličkog guanidin-3,5 "monofosfata (cGMP) sekundarnog glasnika u ciljnoj ćeliji. Povećanje koncentracije cGMP u ciljanim ćelijama pokreće kaskadu biokemijskih reakcija koje mijenjaju njihovo funkcionalno stanje, na primjer, opuštanje stanica glatkih mišića plovila.
Receptori tipa III uključuju receptore koji pod utjecajem endogenih biološki aktivnih tvari - neurotransmitera, osiguravaju prolazak odgovarajućih iona kroz staničnu membranu, što dovodi do promjene njegovog (membranskog) električnog naboja (potencijala).
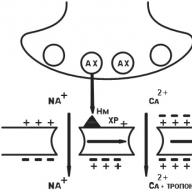
III tina receptori u svojoj strukturi predstavljaju kanal koji prodire kroz lipidni dvoslojni ćelijski membranski oblik, formiran od više poli-cevnih jedinica (Sl. 1.10). Na primjer, nikotinski (H) receptor je kanal promjera 8 nm, formiran od pet polipeptidnih podjedinica (a - dvije, β, γ, d) (vidi Sliku 1.10). Kada neurotransmiter acetilkolin uđe u interakciju s dijelom (domenom) koji strši iznad površine ćelijske membrane, α-podjedinica receptora, njegova struktura se mijenja i otvara se središnji kanal, kroz koji ioni Na + ulaze u ciljanu ćeliju prema koncentracijskom gradijentu, što povlači za sobom promjenu njegove funkcionalne aktivnost. Pored H-kolinergičkih receptora, receptorima tipa III pripadaju i receptori za gama-aminobuternu kiselinu i ekscitacijske aminokiseline.
Sl. 1.10. Dijagram strukture receptora tipa 111:
a je dijagram kola; b - transmembranski ionski kanal (u kontekstu); c - transmembranski ionski kanal (pogled odozgo); / - kanal u neaktivnom (zatvorenom) stanju; 2 - kanal u aktivnom (otvorenom) stanju; a. polipeptidne podjedinice β, γ, d - kanala
Receptori tipa IV uključuju unutarćelijske i nuklearne receptore. Biološki aktivne tvari koje su u interakciji s ovom vrstom receptora su lipofilna (lako topljiva u mastima) spojevi, stoga lako prodiru u staničnu membranu i dopiru do njihovih unutarćelijskih receptora. Intraćelijski receptori uključuju receptore za hormone, kao i druge biološki aktivne tvari.
Mehanizam interakcije hormona s unutarćelijskim receptorima je prilično komplikovan, međutim, on se shematski može prikazati na sljedeći način. Po strukturi, unutarćelijski receptor za hormone je polipeptid koji se sastoji od nekoliko funkcionalnih jedinica - domena. U nedostatku hormona, receptor je neaktivan zbog činjenice da njegov aktivni centar blokira specijalizirani protein - takozvani protein toplotnog udara. U slučaju kada se hormon "približava" receptoru, protein toplotnog udara "odlazi" iz aktivnog centra receptora, s kojim hormon stupa u interakciju (Sl. 1.11).
Nastali receptorsko-hormonski kompleks prodire u jezgru stanice gdje se veže na hormone osjetljive elemente koji se nalaze na DNK (deoksiribonukleinska kiselina; DNA je makromolekula koja se sastoji od zasebnih segmenata - nukleotida, uz pomoć kojih se nasljedne informacije kodiraju u genima; gen - komad DNK koji kontrolira stvaranje jednog strogo definiranog proteina
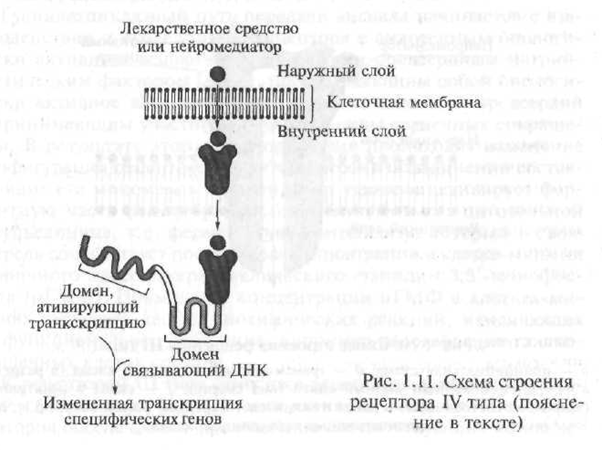
Sl. 1.11. Shema strukture receptora tipa IV (objašnjenje u tekstu)
ka). Kao rezultat ove interakcije započinje proces transkripcije gena - proces prijenosa informacija sadržanih u genetskom kodu iz molekule DNA u molekulu informacijske RNA (mRNA, syn: matrica RNA - mRNA). Transkripcija je prvi korak u stvaranju proteina u ćeliji. Rezultirajuća mRNA. napušta ćelijsko jezgro i prelazi na ribosome - unutarćelijske organele odgovorne za sintezu proteina u ćeliji. U posebnoj medicinskoj literaturi receptore, čije aktiviranje uzrokuje proces transkripcije gena, nazivaju se genetski aktivnim receptorima.
U pravilu, odgovor ciljnih stanica na pobuđivanje gensko aktivnih receptora razvija se polagano, što je od vrlo važne kliničke važnosti.
Prvo, odgovor ciljnih ćelija kasni s vremenom, jer zahteva sintezu novih proteina, što obično traje 20-30 minuta, tj. hormoni, koji aktiviraju receptore tipa IV, nisu u stanju promijeniti patološko stanje u roku od nekoliko minuta, na primjer, odmah zaustaviti napad bronhijalne astme.
Drugo, učinak izazvan ekscitacijom gen-aktivnih receptora prilično je dug i može trajati nekoliko sati ili čak dana, dok se sadržaj lijeka koji su aktivirali ove receptore u krvnoj plazmi smanjuje na nulu mnogo brže. Trajanje efekta u ovom slučaju nastaje usporavanjem biohemijske cirkulacije enzima i proteina koji se sintetišu kao rezultat transkripcije gena. Klinički se to izražava činjenicom da ne postoji povezanost (odnos) između sadržaja plazme određene grupe lijekova i njihovog terapijskog učinka.
Endogene biološki aktivne supstance koje svoj biološki efekat ostvaruju interakcijom sa citosolnim gensko-aktivnim receptorima uključuju steroidne hormone (gluko- i mineralokortikosteroide, polni hormoni), štitnjačne hormone (trijodtironin, tetraiodotironin) i vitamin D rastvorljiv u mastima.
Pored citosolnih gensko-aktivnih receptora, postoje i druge grupe citosolnih receptora, čije pobuđivanje podrazumeva promenu funkcionalne aktivnosti ciljnih ćelija, a ne zbog transkripcije gena.
Takvi receptori uključuju, na primjer, citosolne receptore za dušični oksid (N0). Dušikov oksid (N0) biološki je aktivna supstanca koja se formira u vaskularnom endotelu. Kao endogena biološki aktivna supstanca, azotni oksid prvi je izoliran iz zečjih luka američkog fiziologa R. F. Furchgott-a 1987. godine i nazvan ga je "endotelnim opuštajućim faktorom - ORF." Dušikov oksid je lipofilni spoj koji lako prodire u staničnu membranu, gdje interakciju sa njegovim specifičnim citosolnim receptorima, što povlači za sobom aktiviranje enzima gvanilat ciklaze. Ovo potonje, pak, potiče sintezu cGMP sekundarnog glasnika, što pokreće kaskadu unutarćelijskih biohemijskih reakcija koje dovode do opuštanja ciljnih ćelija, vaskularnih ćelija glatkih mišića.
Tako se trenutno razlikuju četiri glavna mehanizma i, prema tome, IV tipovi receptora, zbog interakcije s kojom su endogene biološki aktivne tvari i / ili njihovi sintetski analozi, tj. Lijekovi mogu utjecati na funkcionalno stanje ciljnih stanica.
Međutim, to ne znači da je broj poznatih receptora za biološki aktivne tvari ograničen na 4. Oni su neizmjerno veći. To je povezano s činjenicom da, istim temeljnim mehanizmom djelovanja, vrlo velik broj endogenih bioloških supstanci različitih kemijskih struktura može utjecati na funkcionalnu aktivnost stanica. Na primjer, neurotransmiteri norepinefrin i histamin, koji se razlikuju po svojoj kemijskoj strukturi i, prema tome, u receptorima s kojima komuniciraju, pobuđuju signal pobude do ciljnih stanica po istom osnovnom mehanizmu - stimulaciji aktivnosti signalnih G proteina, tj. oboje komuniciraju sa receptorima tipa I.
Stoga su svi trenutno poznati receptori klasificirani ne samo na karakteristikama prenošenja signala unutar unutarćelijskih struktura ciljnih ćelija, već i na imenima onih endogenih biološki aktivnih tvari s kojima posebno interaktivno djeluju.
Treba napomenuti da su receptori dobili svoje ime uzimajući u obzir nazive endogenih biološki aktivnih supstanci s kojima stupaju u interakciju, mnogo prije nego su postali poznati mehanizmi prijenosa signala na ciljne stanice.
Endogene biološki aktivne tvari koje svoj učinak ostvaruju interakcijom sa svojim specifičnim receptorima uključuju neurotransmitere (acetilkolin, norepinefrin, dopamin, histamin, serotonin itd.), Hormone, biološki aktivne tvari tkivnog porijekla - autokaidi (prostaglandini, tromboksani, leukotriene, bradikinin, angiotenzin i dr.). U posebnoj medicinskoj literaturi, sve ove supstance često se kombinuju pod terminom "ligandi" (od lat. Ligo - da se vežu, to jest, tvar koja se može vezati za receptor).
Tako receptori dobivaju ime po nazivu njihovih specifičnih liganda. Na primjer, receptori za neurotransmiter dopamin nazivaju se dopamin, hormon inzulina - inzulin, leukotrien augokaid - leukotrien, itd.
Ubuduće će se u tekstu udžbenika, kako bi se izbjegla zabuna u pogledu mehanizma prijenosa signala od receptora do unutarćelijskih formacija, koristiti termin "receptorski tip", a kada je u pitanju ime receptora zbog liganda koji s njim komunicira , upotrijebit će se termin "vrsta receptora".
U pravilu se mnogi receptori iste vrste dijele na više podtipova, na primjer, adrenergički receptori se dijele na a- i β-adrenergičke receptore, holinergički receptori na M- i N-holinergičke receptore itd. U većini slučajeva podvrste se takođe dele na manje grupe: β1 - i β2-adrenoreceptori, N n - i N m -holinergički receptori itd.
Identificiranje podvrsta receptora i proučavanje mehanizama pomoću kojih endogene biološki aktivne tvari komuniciraju s njima vrlo je važno za modernu farmakologiju, jer omogućava stvaranje lijekova koji stupaju u interakciju sa strogo definiranom podvrstaču receptora. Tako je, na primjer, podjela β-adrenergičkih receptora na β 1 \u200b\u200b(uglavnom lokaliziran na staničnoj membrani srčanih ćelija) i β 2 (lokaliziran, na primjer, na staničnim membranama glatkih mišićnih ćelija bronha) omogućio stvaranje lijekova koji selektivno djeluju na srčani mišić (β-adrenostimulansi ) - lijek nonalazin, i selektivno djeluje na glatke mišiće bronha (β 2 -adrenostimulansi) - lijek salbutamol, itd.
Treba napomenuti da organi i tkiva u tijelu ne sadrže stalan broj receptora i / ili njihovih podvrsta, tj. ona je varijabilna. I patološki procesi i lijekovi mogu promijeniti broj receptora u nekom organu.
Na primjer, koronarnu bolest srca prati porast broja 3-adrenergičkih receptora u srčanom mišiću, a kod bolesnika s hipertenzijom povećava se broj a-i β-adrenergičkih receptora. Antidepresiv imipramin, uz duže korištenje, smanjuje količinu β-adrenergičkih receptora u moždanom tkivu. Postoji dosta takvih primjera.
Afinitet endogenih (proizvedenih u tijelu) neurotransmitera ili lijekova za receptore karakteriziran je izrazom "afinitet", a brzina i snaga njihovog vezanja na receptore naznačeni su terminom "afinitet".
Naravno, interakcija lijekova s \u200b\u200breceptorom nije sama sebi svrha, već bi trebala dovesti do neke promjene u radu organa ili tkiva tijela.
Takva promjena, odnosno reakcija, koja odgovara funkcionalnom značaju ovog receptora, naziva se unutarnjom aktivnošću lijekova.
Lijekovi s unutarnjim djelovanjem i afinitetom receptora su agonisti, tj. ponašaju se poput endogenih biološki aktivnih tvari.
Na primjer, fenilefrin, stimulator a-adrenergičkih receptora, ima utjecaj na arteriole slične neurotransmiteru norepinefrinu, tj. on je agonist a-adrenergičkih receptora. U specijalnoj medicinskoj literaturi, uz pojam „agonist“, ponekad se upotrebljava i termin „receptor stimulanse“ ili „mimetik“, na primjer, adrenomimetik, tj. Lijekovi koji stimulišu adrenergičke receptore.
Lijekovi koji imaju afinitet receptora, ali inhibiraju egzogene i endogene agoniste da stupaju u interakciju s receptorom, nazivaju se antagonisti.
Na primjer, atropinski blokator M-kolinergičkih receptora interferira sa interakcijom acetilkolin neurotransmitera M-holinergičkih receptora; blokator (propranolol J-adrenergički receptori, blokirajući β2 -adrenostrukture pluća, inhibira stimulirajući učinak β2 -adrenostimulatora salbutamola na njih, tj. atropin i propranolol su antagonisti odgovarajućih receptora.
U specijaliziranoj medicinskoj literaturi, uz pojam „antagonist“, pojam „blokatori receptora“ ili „litici“, na primjer, antiholinergičari, tj. lijekovi koji blokiraju holinergičke receptore.
Shematski, interakcija receptora sa agonistima i antagonistima prikazana je na Sl. 1.12.
Agonisti mogu ispoljavati i izravno i indirektno, tj. indirektno djelovanje.

Sl. 1.12. Šema interakcije receptora sa agonistom (a) i antagonistom (6) sa receptorom (objašnjenje u tekstu)
Na primjer, agonist opioidnih receptora - lijek morfij - ostvaruje svoj utjecaj direktnim stimuliranjem (mu) p-, (kappa) k- i (delta) 8-opioidnih receptora, tj. vrši direktno stimulativan učinak na njih, dok simpatomimetički efedrin ostvaruje svoje ćelijske efekte indirektno ili indirektno stimulišući a- i β-adrenergičke receptore: pomaže pri uklanjanju neurotransmitera norepinefrina iz presinaptičkih živčanih završetaka, inhibira njegovo vraćanje živčanim završetcima i povećava osjetljivost a- i p- adrenoreceptora do norepinefrina i epinefrina i potiče oslobađanje adrenalina iz kore nadbubrežne kore. Dakle, efedrin svoje farmakološke efekte ostvaruje ne direktno, već kroz neurotransmiter, dok direktno ne komunicira s receptorom.
Antagonisti, kao i agonisti receptora, mogu ostvariti svoje farmakološke učinke direktno ili indirektno, blokirajući odgovarajuće receptore.
Na primjer, želučani blokator histaminskih receptora H3, ranitidin, izravno komunicira sa receptorima histamina želuca H2, inhibira bazalnu sekreciju klorovodične kiseline, inhibirajući interakciju s endogenim receptorima za neurotransmiter histamina, tj. direktno blokira receptor, dok simpatolitički rezerpin, ulazeći u tijelo, poput simpatomimističkog efedrina „izbacuje“ neurotransmiter norepinefrin iz presinaptičkih završetaka, istovremeno blokirajući sintezu novih „porcija“ norepinefrina, što dovodi do brzog iscrpljivanja rezervi medijatora u presinaptičkim završecima. Kao rezultat, nastaje stanje kada a-adrenergički receptori žila "ne djeluju" zbog nedostatka njihovog fiziološkog stimulatora noradrenapina. Možemo reći da simpatolitički rezerpin uzrokuje funkcionalnu, indirektnu blokadu a-adrenergičkih receptora.
Neki lijekovi kombiniraju svojstva agonista i antagonista, tj. pod određenim uvjetima isti receptori pobuđuju ili blokiraju. U onim slučajevima kada stimulativna komponenta prevladava u farmakološkom učinku lijeka, kažu da je lijek djelomični ili djelomični agonist.
Mehanizam djelovanja djelomičnih ili djelomičnih agonista je taj da su ti lijekovi, koji se približavaju receptoru, fiksirani na njemu. Međutim, zbog osobitosti njihove kemijske strukture, oni ih uzbuđuju na takav način da ne uzrokuju potpuni, već samo djelomični stimulirajući odgovor. Istovremeno, oni ometaju interakciju drugih lekova i / ili odgovarajućeg neurotransmitera sa ovim receptorima.
Na primjer, nalorfin, koji se koristi za liječenje predoziranja opojnih analgetika, djelomični je agonist. Nalorfin, „idući“ na opioidni receptor, uklanja morfij iz njegove veze, odnosno zaustavlja agonistički učinak morfija na receptor. Dodirujući receptor, nalorfin u manjoj mjeri ima na njega stimulativan učinak, koji nije u stanju suzbiti aktivnost respiratornog centra, tj. ima djelomični agonistički učinak na opioidni receptor, paralelno s njim blokira ga i morfij, iskazujući tako antagonistički učinak u odnosu na morfij. Nedostatak nalorfina je taj da u slučaju predoziranja pojačava njegov podražajni učinak na receptor i on inhibira respiratorni centar, kao i morfij.
U onim slučajevima kada učinak blokiranja prevladava u farmakološkom učinku lijekova, kažu da je lijek antagonist sa vlastitom ili unutrašnjom aktivnošću.
Osobitosti mehanizma djelovanja antagonista s njihovom vlastitom aktivnošću mogu se razmotriti na primjeru β -adrenoblokatora acebutalola. Acebutalol spada u skupinu β 1 -adrenergijskih blokatora sa vlastitim unutarnjim simpatomimetičkim djelovanjem, tj. kombinira svojstva blokatora (litik) i stimulanse (mimetik) β 1 -adrenoreceptora.
Mehanizam djelovanja acebutalola nastaje zbog osobitosti njegove kemijske strukture, zbog koje lijek s jedne strane blokira miokardni β -adrenoreceptori, a s druge strane stimulira njihovu aktivnost, simulirajući fiziološki učinak posrednika - kateholamina (vidi T. 1. str. 170).
Svi lijekovi prema karakteristikama njihova djelovanja mogu se podijeliti u dvije velike grupe - sa specifičnim i nespecifičnim efektima.
Lijekovi koji imaju nespecifičan učinak uključuju lijekove koji imaju širok spektar farmakološkog djelovanja i različite točke primjene djelovanja. U ovu grupu spadaju vitamini, biogeni stimulansi, antioksidanti, adaptogenovi itd.
Lijekovi sa specifičnim učinkom uključuju lijekove za koje je karakteristično da svojstva agonista ili antagonista odgovarajućih receptora, tj. osnova njihovog mehanizma djelovanja je sposobnost interakcije s određenim receptorima za njih.
Međutim, treba napomenuti da farmakološki učinak lijekova koji imaju specifičan učinak na bilo koje receptore neće uvijek biti visoko specijaliziran, na primjer, smanjenje brzine otkucaja srca (HR) ili suzbijanje izlučivanja želudačnog soka. Ukupni farmakološki učinak mnogih lijekova ovisit će o tome koliko su i u kojim organima i tkivima u tijelu specifične tvorbe receptora za ovaj lijek.
Na primjer, β-adrenergički receptori smješteni su u srčanom mišiću, vaskularnom zidu, bronhima, maternici, masnom tkivu, skeletnim mišićima itd. Kao rezultat toga, lijekovi koji stimulišu β-adrenergičke receptore do određene će mjere uzrokovati povećanje snage i učestalosti kontrakcija srca, širenje krvnih sudova, bronha i niži tonus maternice, tj. Imati sistemski učinak na tijelo.
Specifični efekti lijekova mogu biti selektivni ili selektivni i, shodno tome, neselektivni ili neselektivni. Selektivnost lijekova određuje se utječe li na sve podvrste bilo kojih receptora ili na njihove posebne podvrste.
Na primjer, neselektivni β-adrenoblokator propranolol blokira i β1-adrenoreceptore smještene u miokardnom tkivu i β2-adrenoreceptore smještene, naročito u plućnom tkivu, tj. Ima neselektivni (neselektivni) učinak na obje podvrste β-adrenergičkih receptora. . budući da selektivni β -adrenergički blokator atenolol u terapijskim dozama blokira samo β 1 miokardijalne adrenergičke receptore, tj. ima selektivni (selektivni) efekat na određeni β-adrenergički receptor.
Razjašnjenje fiziološke uloge i lokalizacija receptora podvrste omogućava stvaranje visoko efikasnih lekova koji selektivno deluju na različite podvrste receptora.
Na primjer, lijekovi koji blokiraju β -adrenergičke receptore široko se koriste u klinici za liječenje koronarne srčane bolesti, arterijske hipertenzije, kao i lijekovi koji stimulišu β2-adrenergičke receptore našli su svoju primjenu u liječenju bronhijalne astme. Lijekovi koji blokiraju H1 histaminske receptore preferirano se propisuju u kliničkoj praksi za sprečavanje i / ili zaustavljanje alergijske reakcijedok su lijekovi koji blokiraju H2 histaminske receptore efikasni u liječenju čira na želucu i dvanaestopalačnom crevu.
Stvaranje lijekova sa selektivnim efektom, velikim je dijelom optimiziralo njihovu kliničku upotrebu, jer su značajno smanjeni nuspojava.
Osnova mehanizma djelovanja lijekova u pravilu je njihova sposobnost pokretanja (pokretanja) složenih biokemijskih i / ili biofizičkih procesa koji u konačnici mijenjaju i / ili optimiziraju funkcionalnu aktivnost ciljne stanice.Ljekovi mogu ostvariti svoj učinak na organe i / ili ciljane ćelije putem: direktne hemijske interakcije; fizičko-hemijske interakcije na staničnoj membrani; djelovanje na specijalizirane enzime; akcije na regulatorne gene; delovanje na specifične receptore.
Neposredna hemijska interakcija lijekova.
Taj mehanizam djelovanja lijekova je prilično rijedak i može se realizirati izvan ćelije, na primjer, u lumenu želuca ili crijeva. Njegova suština leži u činjenici da lijekovi ulaze u direktnu hemijsku reakciju s molekulima i / ili ionima koji se u tijelu formiraju u normalnom ili patološkom stanju. Primjer izravne kemijske interakcije je kemijska reakcija neutralizacije klorovodične kiseline želuca prilikom uzimanja antacidnih lijekova.
Fizičko-hemijska interakcija lijekova na staničnu membranu.
Jedna od glavnih funkcija citoplazmatske membrane je provođenje ionske razmjene između citoplazme i izvanćelijskog okruženja. Transmembranska ionska razmjena može se odvijati i kroz posebne transmembranske ionske kanale koji ovise o naponu - natrijum, kalijum, kalcijum, hlor itd. Neki lijekovi, dospijevajući do ćelijske membrane, stupaju u interakciju sa tim kanalima i mijenjaju njihovu funkcionalnu aktivnost. Tako, na primjer, antiaritmično djelovanje lijeka klase IA - kinidin - temelji se na njegovoj sposobnosti da blokira prolazak Na + jona kroz transmembranske natrijumske kanale.
Učinak lijekova na specijalizovane enzime.
Relativno mala količina lijekova ostvaruje njegov farmakološki učinak mijenjanjem aktivnosti nekih specijaliziranih ćelijskih enzima. Zovu se lijekovi koji povećavaju aktivnost staničnih enzima induktorienzimi. Takvim djelovanjem posjeduju, na primjer, tablete za spavanje i antikonvulzivni lijek fenobarbital, koji značajno pojačava aktivnost mikrosomalnih enzima jetre. Biološki značaj ovog efekta fenobarbitala i povezanih lijekova bit će obrađen u daljnjem tekstu.
Nazivaju se lijekovi koji inhibiraju djelovanje specijaliziranih enzima inhibitorienzimi. Tako, na primjer, antidepresiv iz skupine inhibitora monoamin oksidaze (MAO), lijek pirindol realizuje svoje antidepresivno dejstvo inhibiranjem aktivnosti enzima MAO u centralnom nervnom sistemu. Sposobnost inhibiranja aktivnosti enzima acetilkolinesteraza osnova je farmakološkog djelovanja antiholinesteraznih lijekova, na primjer fizostigmina. Poznato je da u fiziološkim uslovima acetilholinesteraza inaktivira (uništava) acetilholin, neurotransmiter koji prenosi ekscitaciju u sinapsama parasimpatičkog nervnog sistema. Fizostigmin, suzbijajući aktivnost acetilholinesteraze, pospješuje nagomilavanje u sinapsama parasimpatičkog sistema neurotransmitera acetilkolina, uslijed čega se povećava ton parasimpatičkog živčanog sustava, što se očituje na sistemskom nivou razvojem bradikardije, snižavanja krvnog tlaka (zglobnog tlaka, lučenja krvi (mokraćnog sustava), lučenja (krvožilnog trakta) (zglobni tlak) (mokraćni trakt), zglobni tlak (mokraćni trakt), zglobni tlak (mokraćni sustav), lučenje krvnih stanica (lučenje krvnih stanica) učenik itd. Lijekovi mogu djelovati reverzibilno i nepovratno s enzimima. Na primjer, lijek enalapril reverzibilno inhibira aktivnost enzima koji pretvara angiotenzin, što posebno povlači za sobom smanjenje krvnog pritiska, dok toksične supstance organofosfor nepovratno inhibiraju aktivnost acetilkolinesteraze. Učinak lijekova na regulatorne gene.Trenutno naučnici pokušavaju stvoriti lijekove koji ostvaruju svoje farmakološke učinke direktno utječući na fiziološku aktivnost regulatornih gena. Ovaj se trend čini posebno obećavajući nakon što je 2000. godine dešifrirana struktura ljudskog genoma. Vjeruje se da će selektivna normalizacija funkcije regulatornih gena pod utjecajem lijekova omogućiti postizanje uspjeha u liječenju mnogih, uključujući i ranije neizlječivih, bolesti. Učinak lekova na receptore.Prije nego što prijeđemo na karakteristike interakcije lijekova s \u200b\u200breceptorima, potrebno je razjasniti što podrazumijevamo pod pojmom "receptor" (od lat. recipio - uzmi, uzmi). Iz tijeka fiziologije poznato je da pojam "receptor" znači visoko specijalizirane formacije koje su u stanju opažati, transformirati i prenositi energiju vanjskog signala u živčani sustav. Ti se receptori nazivaju senzorno(od lat. sensus - osjećanje, osjet, opažanje). Senzorni receptori uključuju receptore organa sluha, vida, mirisa, ukusa, dodira itd. Senzorni receptori ovih organa pripadaju takozvanim eksteroreceptorima. Ako je prisutnost osjetilnih organa koji reagiraju na vanjske podražaje iritacije poznata od davnina, tada je prisutnost senzornih receptora unutar tijela bila dovedena u pitanje do sredine 19. stoljeća. Prvi put je ruski fiziolog I.F. Sion, koji je 1866. godine pokazao pad krvnog pritiska zbog iritacije aorte u pokusu sa kunićima. Ovo je otkriće dovelo do pretraživanja i proučavanja receptora smještenih unutar tijela, a ti receptori su sami nazvani interreceptore. Započetak XX veka otkriven je dovoljan broj senzornih interoreceptora i dokazana njihova važna uloga u regulaciji fizioloških funkcija tijela. Godine 1905. J. Langley je dokazao da se, kada se lijek nanosi na staničnu membranu, razvija farmakološki učinak ako se primjenjuje samo na određeno područje nje. Pored toga, ovo mesto čini samo mali deo ukupne površine ćelijske površine. Ovo opažanje je omogućilo J. Langleyju da zaključi da na staničnoj membrani postoje specijalizirana mjesta receptora koja su u interakciji s lijekovima. Međutim, prioritet u stvaranju recepcijske teorije djelovanja lijekova pripada njemačkom fiziologu P. Ehrlichu, koji je 1906. prvi put uveo termin "receptor" i formulirao postulat "lijek ne djeluje ako nije fiksiran na staničnoj membrani". Prema teoriji P. Ehrlicha, molekula lijeka ima dvije funkcionalno aktivne skupine, od kojih jedna osigurava njegovo fiksaciju na staničnoj površini u regiji receptora za lijek, a druga funkcionalna skupina djeluje u interakciji s receptorom i pokreće složen lanac biohemijskih reakcija koje mijenjaju njegovu (staničnu) fiziološku aktivnost . Dakle, već početkom 20. stoljeća. postalo je očigledno da postoje najmanje dvije klase interoreceptora: senzorni receptoriprenošenje informacija o stanju unutrašnjih organa i tjelesnih tkiva u središnjem živčanom sustavu; ćelijski receptorikoji reagiraju na lijekove koji mijenjaju funkcionalnu aktivnost ciljnih stanica.<>Odmah treba napomenuti da se u budućnosti u tekstu udžbenika, kako bi se izbjegla konfuzija u terminologiji, receptori za lijekove i biološki aktivne tvari, tj. ćelijski ili citoreceptoryoznačit će se terminom "receptor", dok će senzorni interoreceptori biti označeni pojmom koji karakterizira njihovu funkcionalnu aktivnost, na primjer, "baroreceptori", "receptori za bol" itd. Otkriće P. Ehrlicha na staničnoj membrani receptora za lijekove poslužilo je kao polazna osnova za razvoj farmakološke znanosti, posebice farmakodinamike, čiji je jedan od glavnih zadataka proučavanje receptorskih mehanizama djelovanja lijekova. Trenutno su otkrivene struktura velikog broja staničnih receptora, značajke interakcije određenih biološki aktivnih spojeva s njima, što je omogućilo, s jedne strane, razumijevanje mehanizma djelovanja poznatih lijekova, a s druge strane, to je osnova za stvaranje novih vrlo učinkovitih lijekova. Naravno, teško je zamisliti da su se tijekom evolucije u ljudskom tijelu formirali receptori za različite sintetičke (hemijski dobivene) lijekove, posebno jer je velika većina lijekova predstavljenih na modernom farmaceutskom tržištu sintetizirana u posljednjih 50 godina ili manje. Dokazano je da je receptorski aparat ćelije vrlo drevna funkcionalno-strukturna formacija. Dakle, a- i b-adrenoreceptori (receptori, čija međusobna interakcija norepinefrina i adrenalina utječu na funkcionalnu aktivnost stanice) nalaze se ne samo u životinjskim ćelijama, već i na staničnoj membrani biljnih stanica, na primjer, u stanicama biljne nittele, gdje su a- i b- adrenoreceptori regulišu kretanje protoplazme (ćelijski sadržaj). Koji su onda receptori za lijekove koje je otkrio P. Ehrlich i zašto s njima interakciju? Trenutno nema sumnje da su takozvani receptori lijekova zapravo receptori za endogene (proizvedene u tijelu) biološki aktivne tvari uključene u regulaciju funkcionalne aktivnosti unutarnjih organa i tjelesnih tkiva. Takvi biološki aktivni spojevi uključuju tvari koje se oslobađaju iz živčanih završetaka u vrijeme prijenosa živčanog signala, kao i hormone, vitamine, aminokiseline itd. Za svaku endogenu biološki aktivnu tvar postoje strogo specifični receptori za nju. Tako, na primjer, biološki aktivna supstanca proizvedena u tijelu, adrenalin, može aktivirati strogo određene a - i b-adrenergički receptori, a glukokortikosteroidi - hormoni nadbubrežne kore - djeluju samo sa glukokortikosteroidnim receptorima koji su za njih strogo specifični. Sintetski lijekovi koji svoj učinak ostvaruju interakcijom s receptorskim aparatom stanice, po svojoj kemijskoj strukturi manje su više slični endogenim biološki aktivnim spojevima koji stupaju u interakciju sa sličnim receptorima. Na primjer, sintetički vazokonstrikcijski lijekovi (koji uzrokuju vazokonstrikciju) lijekovi fenilefrin su po svojoj kemijskoj strukturi bliski endogenoj biološki aktivnoj tvari norepinefrinu, stoga, poput norepinefrina, ima sposobnost stimulacije a-adrenoreceptora. Ponekad, zbog osobitosti njihove kemijske strukture, lijekovi mogu komunicirati ne sa samim receptorom, već sa susjednim dijelom stanične membrane. Budući da u ovom slučaju, lijek djeluje ne sa samim receptorom, već sa susjednim dijelom stanične membrane, oni ne govore o uzbudljivom ili blokirajućem efektu na receptor, već allostbogati(s grčkog allos - još jedan, drugačiji) učinak, odnosno efekat. Kao rezultat toga, može doći do promjene u strukturi membrane pored receptora i pojedinačnih komponenti samog receptora, što može podrazumijevati promjenu osjetljivosti receptora na biološki aktivnu tvar specifičnu za njega. U slučajevima kada se povećava osjetljivost receptora na biološki aktivnu tvar, razgovarajte senzibilizacija(od lat. sensus - osjećaj) ili otprilike senzibilizacija(od lat. sensibilis - osjetljivost) receptora, a u onim slučajevima kad osjetljivost receptora opada, razgovarajte o tome desesenzibilizacijareceptor. Posebnost alosteričnog efekta leži u činjenici da lijekovi koji imaju ovakav mehanizam djelovanja ne utječu izravno na prijenos živčanog impulsa, već ga modificiraju u željenom smjeru. Na primjer, mehanizam djelovanja anksiolitika (lijekovi protiv anksioznosti; sinonimi: sredstva za smirenje), po svojoj kemijskoj strukturi koji su derivati \u200b\u200bbenzodiazepina, zasnovan je na fenomenu alosternog pobuđivanja postinaptičkih receptora benzodiazepina. Pobuđenje potonjih, pak, potiče aktiviranje inhibicijskih postsinaptičkih receptora gama-aminobuterne kiseline (GABA receptora), što se klinički očituje eliminacijom simptoma neurotičnih bolesti kao što su anksioznost, anksioznost, strah, itd. Receptori, u interakciji s kojima, biološki aktivna tvar ili lijek na bilo koji način mijenja funkcionalno stanje ciljane ćelije, nazivaju se specifične. Pored specifičnih receptora, tzv nespecifičnoza receptore za lijekove. U specijaliziranoj medicinskoj literaturi ovi receptori se nazivaju i "mjesto gubitka" lijekova. Kontaktom s takvim receptorima lijekovi nemaju nikakvog biološkog učinka, ali sami postaju biološki neaktivni. Primjer ove vrste receptora mogu poslužiti kao receptori smješteni na proteinima plazme, posebno na proteinima topivim u vodi - albuminu. Struktura receptora je prilično složena, ali većina je proteinskih makromolekula ili glikoproteina koji mogu uključivati \u200b\u200bione, lipide, nukleinske kiseline itd. Receptor tj. proteinu makromolekulu koja ga tvori odlikuje se specifičnim, specifičnim za svaki receptor, prostornim rasporedom njegovih hemijskih grupa. Proteinska makromolekula koja tvori receptor može se integrirati (uroniti) u lipidni dvosloj citoplazmatske membrane ili lokalizirati unutar ćelije. Glavna funkcija ćelijskog receptora je da „prepozna“ hemijski signal koji mu se prenosi putem endogene biološki aktivne supstance i / ili lekova i transformiše ga u odgovarajući biohemijski i / ili biofizički odgovor ćelije. Ranije se vjerovalo da lijekovi ili endogene biološki aktivne tvari djeluju u interakciji s receptorima tipa "ključ i bravica", tj. receptor ima takvu strukturu koja omogućava lijeku da nađe "vaš" receptor, poveže se s njim i, po njemu, "uključi" i "isključi". Međutim, razvojem medicinske nauke postalo je očito da to nije sasvim istina. Trenutno su molekularni procesi pretvorbe izvanstaničnih signala u unutarćelijsku, regulirajući staničnu funkciju, već prilično proučeni. mehanizmi koji rezultiraju djelovanjem interakcije endogenih biološki aktivnih tvari ili lijekova s \u200b\u200breceptorima. Kod interakcije s receptorom endogene biološki aktivne tvari i / ili aktivnog lijeka poput njega konformacija- prostorna promjena u obliku proteinske makromolekule, što je pokretač različitih procesa unutarćelijskih procesa koji određuju odgovor ciljane stanice na posrednika i / ili lijeka. Na primjer, aktiviranje b2-adrenoreceptora glatkih mišića bronha pod utjecajem b2 -adrenostimulatora fenoterola dovodi do povećanja aktivnosti enzima adenilat ciklaze, što pridonosi nakupljanju cikličkog adenosinofonofata (cAMP) u stanici i, kao rezultat, ćelijskoj relaksaciji. U opštem biološkom smislu, ćelijski receptori se mogu smatrati strogo specijalizovanim „čulnim organima“ ćelija, kroz koji oni opažaju „informacije“ koje potiču, na primer, iz centralnog nervnog sistema i / ili endokrinog sistema. Uprkos važnoj ulozi receptora, receptori zauzimaju samo neznatan dio stanične membrane. Na primjer, M-holinergički receptorski aparat stanice zauzima ne više od 1/6 000 svoje površinske površine. Proučavanje karakteristika interakcije lijekova s \u200b\u200breceptorom, s jedne strane, omogućava nam razumijevanje osnova molekularnog mehanizma njegovog djelovanja, a s druge strane, daje podatke o tome koje bi promjene trebale biti učinjene u strukturi lijekova kako bi se povećala njegova sposobnost interakcije s ovim receptorom, tj. .mogućava ciljanu sintezu novih vrlo efikasnih lijekova. U fiziološkim uvjetima različiti ćelijski receptori ne funkcioniraju neovisno, ali su u stalnoj interakciji jedni s drugima, čime reguliraju specifičnu aktivnost stanice. Na primjer, aktiviranje srčanih b-adrenergičkih receptora endogenim norepinefrinom uzrokuje, posebno, porast broja srčanih kontrakcija i aktiviranje M-holinergičkih receptora srčanih stanica endogenim acetilkolinom, naprotiv, uzrokuje smanjenje broja srčanih kontrakcija. Otkrivanjem pre- i postsinaptičkih receptora velik je doprinos razumijevanju mehanizama djelovanja lijekova. Sinap(s grčkog sinopsis - veza) je specijalizirana kontaktna zona između živčanih ćelija ili drugih uzbudljivih struktura tijela koja osigurava prijenos dolaznih informacija i očuvanje njenog informacijskog značenja. Proučavanje strukture i funkcionalne uloge sinapsi započelo je krajem 19. vijeka. nakon što je španski histolog S. Ramon-i-Cajal (S. Ramon kod Cajala) predložio prisustvo specijalizovanog prenosnog sistema u centralnom nervnom sistemu. Sinapsi su dobili svoje ime 1897. godine, kada je engleski fiziolog Ch. Sherrington predložio ovaj termin da se odnosi na područje dodira između nervnih ćelija. Trenutno postoje tri vrste sinapsi: 1) "električne" sinapse u koje se informacije prenose prijenosom električnog signala iz pred-sinaptičke membrane. Ova vrsta sinapse se zove efaps(s grčkog ephapsis - tijesan kontakt); 2) "hemijske" sinapse u kojima se informacije prenose putem posebnih biološki aktivnih supstanci - neurotransmiteri(s grčkog neuron - nerv i lat. posrednik - posrednik; sinonimi: medijator); 3) „mješovite“ sinapse u kojima se informacije prenose i hemijski i električno. Farmakološki učinci ogromne većine lijekova koji utječu na funkcije sinapsi ostvaruju se svojim učinkom na određeni stadij prijenosa signala u hemijskim sinapsama, tj. u sinapsama druge vrste. Po pravilu, hemijske sinapse klasificiraju neurotransmiteri koji u njima prenose živčane impulse kako slijedi: sinapse u kojima acetilholin djeluje kao posrednik nazivaju se kolinergičkim; sinapse u kojima adrenalin i norepinefrin djeluju kao posrednik nazivaju se adrenergički; sinapse u kojima ATP i adenozin djeluju kao posrednik nazivaju se purinergički; sinapse u kojima gama-aminobuterna kiselina djeluje kao posrednik nazivaju se GABA-ergičkim itd. Struktura sinapse je trenutno dobro razumljiva. Sinaps se sastoji od presinaptičkog procesa nervne ćelije (akson-kraj) i aparata za prijem „signala“ koji se nalazi na membrani efektorske („izvršne“) ćelije.
Farmakodinamika - farmakološki učinci, mehanizmi djelovanja, lokalizacija djelovanja, vrste djelovanja lijekova.
Farmakološki efekti ljekovita supstanca - promjene u radu organa, tjelesnih sistema koji uzrokuju ovu supstancu (na primjer, pojačane kontrakcije srca, snižavanje krvnog pritiska, poticanje mentalne aktivnosti, uklanjanje straha i napetosti itd.).
Svaka tvar izaziva niz farmakoloških učinaka karakterističnih za nju. U svakom se slučaju koriste samo određeni učinci ljekovite tvari koji su definirani kao glavni efekti. Pozvani su preostali (neiskorišteni, nepoželjni) farmakološki učinci kolateralna.
Mehanizmi delovanja lijekovi - načini na koje tvari izazivaju farmakološke učinke vrlo su raznoliki. Glavne mogućnosti mehanizama djelovanja uključuju akcije na:
- specifični receptori;
- enzimi;
- jonski kanali;
- transportni sistemi.
Većina lekova deluje specifični receptori. Ovi receptori su najčešće predstavljeni funkcionalno aktivnim molekulama proteina, čija interakcija potiče biohemijske reakcije koje dovode do farmakoloških učinaka.
Postoje specifični receptori povezani sa staničnim membranama (membranom) i intracelularni receptori (citoplazmatski, nuklearni).
Membranski receptori (receptori citoplazmatske membrane) dijele se na:
- receptore koji su direktno povezani sa ionskim kanalima;
- receptore direktno konjugirane na enzime;
- receptore koji stupaju u interakciju sa G-proteinima.
Za receptore direktno povezane na ionske kanale, uključuju, posebno, Ν-kolinergičke receptore i GABA A-receptore.
Kada se stimulišu Ν-kolinergički receptori (nikotin-osjetljivi holinergički receptori), natrijumski kanali koji su direktno povezani s njima se otvaraju. Stimulacija Ν-kolinergičkih receptora dovodi do otkrića Na + kanala, ulaska Na + iona u ćeliju, depolarizacije ćelijske membrane i uzbudljivog efekta.
GABA A receptori su direktno povezani s klorovim kanalima. Stimulacija GABA A receptora dovodi do otkrića Cl - kanala, ulaska Cl - jona, hiperpolarizacije ćelijske membrane i inhibitora.
Za receptore koji su direktno povezani sa enzimimauključuju, posebno, receptore za inzulin koji su direktno konjugirani na tirozin kinazu.
G-proteinski receptori - M-holinergički receptori (na muskarinske osjetljive holinergičke receptore), adrenergički receptori, dopaminski receptori, opioidni receptori itd.
G-proteini, tj. Proteini koji vezuju GTP, lokalizirani su u staničnoj membrani i sastoje se od α-, β- i γ-podjedinica. Kada lijek djeluje u interakciji s receptorom, α-podjedinica G-proteina veže se na GTP (GTP) i djeluje na enzime ili ionske kanale.
Jedan receptor u interakciji je s nekoliko G-proteina, a svaki kompleks α-podjedinice G-proteina s GTP djeluje na više molekula enzima ili na nekoliko ionskih kanala. Tako se implementira mehanizam pojačala (pojačala): kada se aktivira jedan receptor, aktivnost mnogih molekula enzima ili mnogih ionskih kanala se mijenja.
Jedan od prvih koji su pronađeni bili su G-proteini povezani sa β-adrenoreceptorima u srcu. Kad se aktivira simpatička inervacija srca, pobuđuju se β -adrenoreceptori; adenilat ciklaza se aktivira putem G-proteina; cAMP nastaje iz ATP-a, aktivira se protein kinaza, pod djelovanjem kojeg se Ca2+ kanali fosforiliraju i otvaraju.
Povećani unos Ca 2+ iona u ćelije sinoatrijskog čvora ubrzava četvrtu fazu akcijskog potencijala, učestalost generisanih impulsa raste - srčane kontrakcije postaju brže.
Otkrivanje Ca2+ kanala u vlaknima radnog miokarda dovodi do povećanja koncentracije Ca 2+ u citoplazmi (unos Ca 2+ potiče oslobađanje Ca 2+ iz sarkoplazmatskog retikuluma). Ioni Ca 2+ vežu se na troponin C (komponenta troponin-tropomiozin); Tako se smanjuje inhibicijski učinak troponin-tropomiozina na interakciju aktina i miozina - pojačavaju se kontrakcije srca (Slika 10).
Sl. 10. Mehanizam povećanja i jačanja srčanih kontrakcija za vrijeme stimulacije β -adrenoreceptora. AC - adenilat ciklaza; PC - protein kinaza; CA - sinoatrijski čvor; Ttm - troponin-tropomiozin.
Kad se aktivira parasimpatička inervacija srca (vagusni živci), pobuduju se M2-kolinergički receptori, a putem G-proteina inhenilat-ciklaza se inhibira - srčane kontrakcije slabe i slabe (uglavnom su atrijske kontrakcije oslabljene, jer je parasimpatička inercija ventrikula relativno slaba).
Dakle, G-proteini mogu imati stimulirajući i inhibirajući učinak na adenilat ciklazu. Stimulirajući G-proteini označeni su kao G s-proteini (stimulišu) i inhibitorni G-proteini (G-proteini (inhibiraju) (Sl. 11).
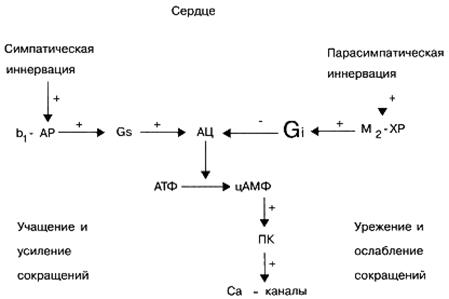
Sl. 11. Mehanizam promjene u učestalosti i jačini srčanih kontrakcija tokom stimulacije simpatičke i parasimpatičke inervacije.
Toksin kolere aktivira C s proteine \u200b\u200b(to dovodi do aktivacije adenilata ciklaze, a kod kolere se manifestuje izlučivanjem tečnosti kroz crevni epitel).
Pertusis toksin aktivira G i proteine.
Kada se M1 -holinergički receptori, M3 -holinergički receptori, α-adrenoreceptori pobude kroz proteine \u200b\u200bG q, aktivira se fosfolipaza C, što doprinosi stvaranju inozitol-1,4,5-trifosfata i diacilglicerola iz fosfatidilinozitol-4,5-difosfata .
Inozitol-1,4,5-trifosfat djeluje na receptore sarkoplazmatske membrane retikuluma osjetljive na njega i potiče oslobađanje Ca 2+ iona iz sarkoplazmatskog retikuluma (Sl. 12). Stimulacijom αι-adrenoreceptora krvnih žila to dovodi do kontrakcije glatkih mišića i sužavanja sudova (slika 13).

Sl. 12. Učinak fosfolipaze C na nivo citoplazme C a2 +.
Osjetljivost receptora na agoniste i broj receptora se stalno mijenjaju. Dakle, nakon stimulacije p1 -adrenoreceptora agonistom β -adrenoreceptora, fosforilat se vrši posebnom receptorskom kinazom, veže se na β-arstinski protein i u tom kompleksu gubi sposobnost interakcije s G-proteinima (desenzibilizacija receptora). Kompleks β-adrenoreceptora sa β-arestininom ćelija apsorbuje endocitozom (internalizacija receptora) i zahvaćaju ga endosomi i lizosomi. U endosomima se β1 -stainski molekuli odvajaju od receptora koji se ponovo integrišu u staničnu membranu; obnavlja se osjetljivost receptora na agoniste (resensitization receptora). U lizosomima dolazi do razaranja molekula receptora (smanjivanje regulacije) (Sl. 14).

Sl. 13. Mehanizam kontrakcije glatkih mišića krvnih žila tokom stimulacije simpatičke inervacije. FLS - fosfolipaza C; FIF 2 - fosfatidilinozitol-4,5-difosfat; Ako je 3 - inozitol-1,4,5-trifosfat; SR - sarkoplazmatski retikulum; KLCM - kinaza lakih lanaca miozina.

Sl. 14. Desenzibilizacija i redukcija β-adrenergičkih receptora.
Za intracelularni receptori uključuju se kortikosteroidni i receptori za polne hormone. Konkretno, glukokortikoidni receptori lokalizirani su u citoplazmi ćelija. Nakon što se glukokortikoid kombinira sa citoplazmatskim receptorima, kompleks glukokortikoida - receptora prodire u jezgro i utječe na ekspresiju različitih gena.
Sposobnost tvari da se vezuju za receptore (sklonost tvari da se vezuju za receptore) označena je terminom " afinitet". U odnosu na iste receptore, afinitet različitih tvari može biti različit. Za karakterizaciju afiniteta koristite indikator strK D je negativni logaritam konstante disocijacije, odnosno koncentracije tvari na kojoj je 50% receptora zauzeto.
Interna aktivnost - sposobnost tvari da stimulišu receptore; određeno od farmakološki učinakpovezan s aktivacijom receptora.
U normalnim uvjetima, ne postoji direktna povezanost između afiniteta i unutrašnje aktivnosti. Supstanca može zauzeti sve receptore i izazvati slab efekat, i obrnuto, supstanca može zauzeti 10% receptora i izazvati maksimalan učinak za ovaj sistem.
Agonisti - supstance s afinitetom i unutrašnjom aktivnošću.
Kompletni agonisti posjeduju afinitet i maksimalnu unutrašnju aktivnost (mogu izazvati maksimalan učinak za neki sistem), čak i ako zauzimaju dio specifičnih receptora.
Djelomični (djelomični) agonisti posjeduju afinitet i manje od maksimalne unutarnje aktivnosti (mogu izazvati samo manje od maksimalnih efekata, čak i ako zauzimaju 100% specifičnih receptora).
Antagonistiposjeduju afinitet, ali ne posjeduju unutrašnju aktivnost i ometaju djelovanje punih ili djelomičnih agonista (istiskuju agoniste iz komunikacije s receptorima).
Ako se djelovanje antagonista eliminira povećanjem doze agonista, takav se antagonizam naziva konkurentnim.
Djelomični agonisti mogu biti antagonisti punih agonista. U nedostatku potpunog agonista, djelomični agonist stimulira receptore i izaziva slab efekat. Kada komunicira s kompletnim agonistom, djelomični agonist zauzima receptore i inhibira djelovanje kompletnog agonista. U ovom slučaju učinak potpunog agonista je oslabljen.
Na primjer, pindolol, djelomični β-adrenoreceptorski agonist, izaziva slabu tahikardiju u nedostatku efekata simpatičke inervacije na srce. Ali s povećanjem tona simpatičke inervacije, pindolol djeluje kao pravi β-blokator i uzrokuje bradikardiju. To je zbog činjenice da djelomični agonist pindolol slabi učinak posrednika norepinefrina, koji je potpuni agonist u odnosu na β-adrenergičke receptore srca.
Agonisti antagonista - tvari koje djeluju različito na podtipove istih receptora: stimulišu neke podvrste receptora, a blokiraju druge. Na primjer, narkotički analgetik nalbufin ima različite učinke na podtipove opioidnih receptora. Nalbuphine stimulira κ receptore (i stoga smanjuje osjetljivost na bol), a µ receptore blokira (i samim tim je manje opasan u smislu ovisnosti o lijekovima).
Primer uticaja supstanci na enzimi može doći do djelovanja antiholinesteraznih sredstava koja blokiraju acetilholinesterazu (enzim koji razgrađuje acetilholin) i na taj način pojačava i produžuje djelovanje acetilkolina.
Poznate ljekovite tvari koje stimulišu ili blokiraju jonski kanali stanične membrane, to jest kanali koji selektivno vode ione Na +, K +, Ca 2+ (natrijum, kalijum, kalcijumski kanali) itd. Na primjer:
Lokalni anestetici blokiraju Na + kanale;
Antiaritmički lijekovi klase I (kinidin, lidokain) blokiraju Na + kanale;
Minoksidil aktivira K + kanale;
Hipoglikemijski agensi iz grupe derivata sulfoniluree blokiraju K + kanale ovisne o ATP-u;
Verapamil, nifedipin blokira Ca 2+ kanala.
Primer uticaja supstanci na transportnih sistema može postojati akcija:
Reserpin (blokira vezikularni unos dopamina i norepinefrina);
Srčani glikozidi (inhibiraju Na + / K + -ATPazu);
Triciklički antidepresivi (blokiraju povratni neuronski unos norepinefrina i serotonina);
Blokatori protonske pumpe (omeprazol itd.).
Mogući su i drugi mehanizmi djelovanja. Na primjer, diuretski manitol povećava diurezu povećanjem osmotskog tlaka u bubrežnim tubulima. Anti-aterosklerotski agens - kolestipol - veže (sekvestrira) žučne kiseline, ometa njihovu apsorpciju u crevima, pa se stoga aktivira stvaranje žučnih kiselina iz holesterola u jetri i smanjuje se nivo holesterola u hepatocitima.
Mehanizmi djelovanja različitih lijekova proučavani su u različitoj mjeri. U procesu njihovog proučavanja, ideje o mehanizmima djelovanja ne samo da mogu postati složenije, već se i značajno promijeniti.
Koncept " lokalizacija akcije„Znači primarno (-e) mjesto (-e) djelovanja određenih ljekovitih tvari. Na primjer, srčani glikozidi djeluju uglavnom na srce.
Na koncept " vrste akcije»Odnosite se za lokalne i opće (resorptivne) akcije, refleksno djelovanje, primarno i sekundarno djelovanje, direktno i indirektno djelovanje.
Primjer lokalne akcije može biti djelovanje lokalnih anestetika.
Većina lijekova ima općeniti (resorptivni) učinak, koji se obično razvija nakon apsorpcije (resorpcije) neke tvari u krvi i njene distribucije u tijelu.
I s lokalnim i s resorptivnim djelovanjem, tvari mogu pobuditi različite osjetljive receptore i izazvati refleksne reakcije.
Glavni efekat ljekovite supstance su njeni efekti, koji se koriste u svakom slučaju. Svi ostali efekti procjenjuju se kao manifestacija nuspojava.
Ljekovite tvari mogu djelovati na određene organe direktna akcija. Osim toga, učinak lijekova može biti indirektan. Na primjer, srčani glikozidi imaju direktan utjecaj na srce, ali, poboljšavajući rad srca, ove tvari povećavaju opskrbu krvlju i funkcije drugih organa (indirektni učinak).