По-голямата част наркотици има терапевтичен ефект чрез промяна на активността на физиологичните системи на клетките, които се произвеждат в организма по време на еволюцията. Под въздействието на лекарствено вещество в организма, като правило, нов вид клетъчна активност не възниква, променя се само скоростта на различни естествени процеси. Инхибирането или възбуждането на физиологичните процеси води до намаляване или засилване на съответните функции на телесните тъкани.
Лекарствата могат да действат върху специфични рецептори, ензими, клетъчни мембрани или директно да взаимодействат с клетъчните вещества. Подробности за механизмите на действие лекарствени вещества изучаван в курс по обща или експериментална фармакология. По-долу предоставяме само някои примери за основните механизми на действие на лекарствата.
Ефект върху специфични рецептори. Рецепторите са макромолекулни структури, които са селективно чувствителни към определени химични съединения. Взаимодействието на химикалите с рецептора води до появата на биохимични и физиологични промени в организма, които се изразяват в определен клиничен ефект.
Лекарствата, които директно възбуждат или повишават функционалната активност на рецепторите, се наричат \u200b\u200bагонисти, а вещества, които инхибират действието на специфични агонисти, се наричат \u200b\u200bантагонисти. Антагонизмът може да бъде конкурентен и неконкурентен. В първия случай лекарственото вещество се конкурира с естествен регулатор (медиатор) за места на свързване в специфични рецептори. Блокада на рецептори, причинена от конкурентен антагонист, може да бъде елиминирана с големи дози агонист или естествен медиатор.
Разнообразие от рецептори са разделени по чувствителност към естествените медиатори и техните антагонисти. Например чувствителните към ацетилхолин рецептори се наричат \u200b\u200bхолинергични, адреналин-чувствителните рецептори се наричат \u200b\u200bадренергични. По отношение на чувствителността към мускарин и никотин холинергичните рецептори са разделени на чувствителни към мускарини (m-холинергични рецептори) и чувствителни към никотин (n-холинергични рецептори). Н-холинергичните рецептори са хетерогенни. Установено е, че разликата им се крие в чувствителността към различни вещества, Има n-холинергични рецептори, разположени в ганглиите на автономната нервна система, и n-холинергични рецептори на набраздения мускул. Известни са различни подтипове на адренергични рецептори, обозначени с гръцките букви α1, α 2, β1, β2.
Изолират се също Н1 и Н2 хистамин, допамин, серотонин, опиоиди и други рецептори.
Ефект върху ензимната активност. Някои лекарства повишават или инхибират активността на специфични ензими. Например, физостигминът и неостигминът намаляват активността на холинестеразата, която унищожава ацетилхолина и дават ефекти, характерни за възбуждането на парасимпатиковата нервна система. Инхибиторите на моноаминооксидазата (ипразид, ниаламид), които предотвратяват разрушаването на адреналина, повишават активността на симпатиковата нервна система. Фенобарбитал и циксорин, увеличавайки активността на чернодробната глюкуронилова трансфераза, намаляват нивото на билирубин в кръвта.
Физико-химичен ефект върху клетъчните мембрани. Активността на клетките на нервната и мускулната система зависи от потока на йони, които определят трансмембранния електрически потенциал. Някои лекарства променят транспорта на йони.
Така че антиаритмични, антиконвулсивни лекарства, лекарства за обща анестезия.
Директно химическо взаимодействие. Лекарствата могат директно да взаимодействат с малки молекули или йони вътре в клетките. Например, етилендиаминтетраоцетна киселина (EDTA) силно свързва оловни йони. Принципът на пряко химическо взаимодействие е в основата на използването на много антидоти за химическо отравяне. Друг пример е неутрализирането на солна киселина с антиациди.
фармакодинамика
Той изучава механизма на действие на лекарствата, както и техните биохимични и физиологични ефекти. Задачите й включват описание на химичните и физичните взаимодействия между лекарството и целевата клетка, както и пълния спектър и тежестта на неговите фармакологични ефекти. Познаването на фармакодинамичните модели ви позволява да изберете правилните лекарства. Фармакодинамичните изследвания осигуряват по-дълбоко разбиране за регулирането на биохимичните и физиологичните процеси в организма (Katzung B.G., 1998; Lawrence D.R. et al., 2002).
Действието на повечето лекарства се медиира от свързването им с макромолекулите на тялото. Промяна във функционалното състояние на тези макромолекули от своя страна задейства верига от биохимични и физиологични реакции, които се превръщат във фармакологичен ефект. Макромолекулите, с които взаимодействат химикалите, се наричат \u200b\u200bрецептори. По този начин, всяка функционално активна макромолекула може да служи като рецептори за лекарства. От това твърдение произтичат няколко важни последици. Първо, с помощта на лекарства можете да промените скоростта на всеки физиологичен процес в организма. Второ, лекарствата променят само естествените физиологични функции на клетката, без да й придават нови свойства.
рецептори
Повечето рецептори са протеини. Това са рецептори на хормони, растежни фактори, медиатори, протеини, участващи в най-важните метаболитни и регулаторни реакции (дихидрофолат редуктаза, ацетилхолинестераза), транспортни протеини (Na +, K + -ATPase), структурни протеини (тубулин). Клетъчните компоненти от различно химично естество, като нуклеинови киселини, с които взаимодействат антитуморни агенти, също могат да действат като рецептори.
Рецепторите на ендогенни регулаторни фактори - хормони, медиатори и др., Имат фармакологично значение. Тези рецептори служат като мишени за много лекарства, като обикновено действат избирателно поради високата специфичност на рецепторите за ендогенни лиганди. Лекарствата, които след свързване с рецептора възпроизвеждат физиологичния ефект на ендогенния лиганд, се наричат \u200b\u200bаганисти или стимуланти. Лекарствата, които не причиняват този ефект, но инхибират свързването на ендогенни лиганди, се наричат \u200b\u200bантагонисти или блокери. Вещества, чийто ефект е по-слабо изразен от ефекта на агонистите, се наричат \u200b\u200bчастични агонисти. Препаратите, стабилизиращи рецептора в неактивирана форма, се класифицират като обратни агонисти.
Структурна и функционална зависимост
Химическата структура на лекарството доста строго определя неговия афинитет към рецепторите и вътрешната активност. Лека промяна в химическата структура може значително да повлияе на фармакологичните свойства.
Синтезът на нови лекарства до голяма степен се основава на това. Тъй като химическата модификация не засяга непременно всички фармакологични свойства еднакво, е възможно да се подобри ефективността и безопасността на лекарството, да се увеличи неговата селективност и да се подобрят фармакокинетичните характеристики. Например, много антагонисти на хормони и медиатори, използвани в клиниката, се синтезират чрез химическа модификация на ендогенни вещества.
Точки за кандидатстване за лекарства
Тъй като ефектът на лекарствата е медииран от рецептори, точката на приложение на лекарството се определя не само от особеностите на неговото разпределение, но и от локализацията на рецепторите, а фармакологичните ефекти зависят от функционалното значение на тези рецептори. Фармакологичните ефекти на лекарствата, чиито рецептори са често срещани в много органи и тъкани, са разнообразни. Ако тези рецептори изпълняват жизненоважна за клетките функция, не само е трудно да се използва лекарството за терапевтични цели, но е и опасно. Независимо от това, такива лекарства могат да бъдат от голямо клинично значение. И така, сърдечните гликозиди, широко използвани при сърдечна недостатъчност, променят транспорта на йони през клетъчната мембрана, от което зависи жизнената активност на клетката. Те имат тесен терапевтичен диапазон и са много токсични. Друг пример са противотуморни средства. Ако рецепторите, с които лекарството взаимодейства, присъстват само на няколко типа диференцирани клетки, неговият ефект е по-селективен. Тези лекарства могат да имат по-малко нежелани реакции, но все пак, тези лекарства могат да бъдат токсични, ако техните рецептори изпълняват жизненоважна функция. Някои биологични отрови (ботулинов токсин и др.) Действат по подобен начин. Освен това, дори ако директният фармакологичен ефект е селективен, последствията от него могат да бъдат по-разнообразни.
Ендогенни рецепторни регулаторни фактори
Терминът рецептор се отнася до всеки макромолекулен компонент на клетка, с който лекарството се свързва. Един от най-важните рецептори за лекарства са клетъчните протеини, които служат като рецептори за ендогенни регулаторни фактори - хормони, растежни фактори, медиатори. Свързвайки се с ендогенния лиганд, рецепторите предават сигнала от него в целевата клетка.
От рецептора сигналът стига до клетъчните мишени (ефекторни протеини) директно или чрез междинни сигнални молекули - протеин-конвертори. Рецепторите, протеиновите конвертори и ефекторните протеини образуват рецептор-ефекторната система. Най-близкият ефекторен протеин във веригата за предаване на сигнал често не е терминален ефектор (пряко засягащ клетъчните функции), а ензим или транспортен протеин, участващ във формирането, транспортирането или инактивирането на втори медиатор - йон или малка молекула. Вторият медиатор от своя страна прехвърля информация към различни вътреклетъчни цели, осигурявайки им едновременна реакция на сигнал от един рецептор.
Рецептори, конвертиращи протеини и ефекторни протеини не само предават информация. Те също координират сигналите от различни лиганди, от една страна, и всички тези сигнали с метаболитни процеси в клетката, от друга.
Действайки като катализатори, рецепторите усилват биологичния сигнал. Поради това важно свойство те служат като отлични мишени за лекарства. Усилвателите на сигнала обаче са не само рецептори с ензимна активност, но всички известни рецептори. В действителност, когато една молекула лиганд се свързва към рецептор, конюгиран с йонния канал, много йони преминават през последния. Същото важи и за рецепторите за стероидни хормони: една хормонална молекула задейства транскрипцията на много копия на мРНК, въз основа на които се синтезират многобройни протеинови молекули.
В зависимост от структурата и механизма на действие, рецепторите на биологично активни вещества се разделят на няколко класа. Броят на тези класове е малък.
Ензимни рецептори
Най-голямата група рецептори с ензимна активност са мембранните рецептори със собствена активност на протеин киназа. Те фосфорилират различни ефекторни протеини, разположени от вътрешната страна на клетъчната мембрана. В резултат на това функцията на тези протеини или тяхното взаимодействие с други протеини се променя.
Има още един клас рецептори с активност на протеин киназа - това са рецептори, конюгирани с протеин кинази. Липсва вътреклетъчен каталитичен домен, но при взаимодействие с агонист те свързват или активират вътреклетъчните протеинкинази на вътрешната повърхност на мембраната. Това са рецептори за невротрофни фактори и антиген-разпознаващи рецептори за Т и В лимфоцити, състоящи се от няколко субединици. Последните също взаимодействат с фосфотирозин фосфати. Функцията на други рецептори, които нямат вътреклетъчен ефектор домен, може да бъде медиирана от някои други ефекторни протеини.
Други рецептори със собствена ензимна активност имат подобна структура. Те включват например рецептори със собствена активност на фосфотирозин фосфатаза: извънклетъчният им домен е подобен в аминокиселинната последователност на адхезионните молекули. За много рецептори със собствена активност на фосфотирозин фосфатазата ендогенните лиганди не са известни. Въпреки това, според генетични и биохимични изследвания, проведени върху различни видове клетки, ензимната активност на тези рецептори играе важна роля. Вътреклетъчният домен на предсърдни рецептори за натриуретичен хормон, други NP, а също и рецептори на гуанилин има собствена активност на гуанилат циклазата и синтезира cGMP, който действа като втори медиатор. Може би има други рецептори със собствена ензимна активност.
Рецептори, свързани с йонни канали
Рецепторите на някои медиатори са пряко свързани с йонни канали, взаимодействащи с лиганд за селективно преминаване на определени йони през клетъчната мембрана (хемочувствителни канали, йонотропни рецепторни канали, йонотропни рецептори).
G-протеинови куплирани рецептори
Това е доста голям клас рецептори, които взаимодействат с ефектори чрез G-протеини (протеини, които използват заместването на гуанин дифосфат (GDF) с гуанин трифосфат (GTP). Те включват рецептори за много биогенни амини, липидни сигнални молекули (по-специално ейкозаноиди) и различни пептиди и протеинови лиганди. Ензимите (аденилат циклаза, фосфолипаза С) и калиевите и калциевите мембранни канали действат като ефектори. Големият брой и важна физиологична роля на рецепторите, съчетани с G-протеините, ги прави отлични. моите цели за лекарства: приблизително половината от всички лекарства, предписани от лекари (с изключение на антибиотици), действат върху тези рецептори.
Клетката може да носи до 20 рецептора на повърхността си, всеки от които избирателно взаимодейства с един или повече видове G-протеини (различават се в различни видове α-субединици). Α-субединицата е в състояние да взаимодейства с един или повече ефекторни протеини, което ви позволява да координирате сигнали от рецептори на различни лиганди, използвайки един G-протеин. От друга страна, един рецептор може да задейства няколко механизма на вътреклетъчно предаване на сигнал, активирайки няколко типа G-протеини и да действа върху различни ефекторни протеини чрез една и съща α-субединица. Такава сложна система на дивергенция и конвергенция на сигналите осигурява гъвкава регулация на клетъчните функции (Ross, 1992).
Вътреклетъчни рецептори
Рецепторите на стероидни и щитовидни хормони, калцитриол и ретиноиди са разтворими вътреклетъчни ДНК-свързващи протеини, които регулират транскрипцията на определени гени (Mangelsdorf et al., 1994). Тези рецептори принадлежат към семейството на чувствителни към лиганда транскрипционни регулатори. Функцията на транскрипционните фактори се регулира от фосфорилиране, взаимодействие с клетъчни протеини, метаболити и други регулаторни компоненти на клетката.
Втори посреднически системи
cAMP. В интегрирането на външни сигнали участват и вторичните посреднически системи. Въпреки че има много повече известни рецепторни и протеинови сигнални молекули от вторите медиатори, последните участват в много пътища в рамките на предаването на клетъчния сигнал. Най-изследваните втори посредници включват cAMP, cGMP, Ca 2+, IF3 (инозитол трифосфат), DAG (диацилглицерол), NO. Тази група хетерогенни съединения непрекъснато расте. Вторите медиатори си взаимодействат пряко (чрез промяна на взаимния метаболизъм) или косвено (като действат върху едни и същи вътреклетъчни цели). Функцията на вторите медиатори, както и регулирането на тяхното формиране (или освобождаване), разцепване и екскреция от клетката, е удобно разгледано с примера на cAMP. Този втори медиатор се синтезира под въздействието на аденилат цитаза при активиране на много рецептори, конюгирани с G-протеини. G s протеинът активира аденилатциклазата, G i протеинът инхибира.
Има поне 10 тъканно-специфични аденилат циклотазни изоформи, които се различават по механизмите на регулиране на активността.
Като правило, сАМР активира протеин кинази А (cAMP-зависими протеина кинази), малка група от свързани протеини. Тези протеинкинази от своя страна фосфорилират не само крайните вътреклетъчни мишени (ензими, транспортни протеини), но и други протеинкинази и други регулаторни протеини. Последните включват например фактори на транскрипция. Те са отговорни за cAMP-медиираното регулиране на генната транскрипция, осигурявайки забавен клетъчен отговор на сигнала. В допълнение към активирането на протеин кинази, cAMP действа директно върху каналите на катионните мембрани, които играят важна роля, по-специално, във функционирането на невроните. По този начин сигналът от cAMP причинява верига от биохимични промени в целевата клетка.
Калций. Друг добре проучен втори медиатор е вътреклетъчен Ca 2+. Йони Ca 2+ навлизат в цитоплазмата по различни начини: през мембранни канали (G-протеин зависими, затворени с напрежение, регулирани от K + или Ca 2+ sim), както и през канали, разположени в специфични зони на ендоплазмения ретикулум и отварящи се под действието на АКО 3 и в скелетния мускул в резултат на мембранната деполяризация. Отстраняването на калций от цитозолната плазма става по два начина: той се абсорбира от ендоплазмения ретикулум или се екскретира от клетката. Са 2+ предава сигнали на много по-голям брой протеини от сАМФ - ензими, участващи в клетъчния метаболизъм, протеин кинази, калций-свързващи протеини. Последните взаимодействат с други крайни и междинни ефекти.
Регулиране на рецепторите
Рецепторите не само контролират физиологичните и биохимичните функции, но също така служат като обекти на регулиране. Това регулиране се осъществява на ниво синтез и разлагане на техните макромолекули, чрез образуване на ковалентни връзки с други молекули, взаимодействие с регулаторни протеини и движение на рецепторите. Конвертиращите протеини и ефекторните протеини също подлежат на регулиране. Регулаторните сигнали могат да идват от вътреклетъчни канали за предаване, активирани чрез стимулиране на самия рецептор (чрез механизъм за обратна връзка), както и от други рецептори (пряко или косвено).
Дългосрочната стимулация на лекарствените рецептори обикновено води до намаляване на реакцията към него - при същата концентрация лекарството предизвиква по-слабо изразен ефект. Това явление, наречено десенсибилизация, рефрактерно, пристрастяване, играе важна роля в клиничната практика: например с продължителна употреба β-адренергичните агонисти за лечение на пациенти с AD тежестта на реакцията към тези лекарства е намалена.
Хомологичната десенсибилизация се отнася само за стимулирани рецептори и е специфична за лиганда. С хетероложна десенсибилизация тежестта на реакцията към други лиганди, чиито рецептори действат по един и същ път на предаване на вътреклетъчен сигнал. В първия случай отрицателната обратна връзка се осигурява от ефекта върху самия рецептор (фосфорилиране, протеолиза, намален синтез), във втория случай, в допълнение към рецептора, той може да засегне и други протеини, участващи във вътреклетъчното предаване на сигнал.
Напротив, ако рецепторите не се стимулират дълго време, тяхната чувствителност към агонисти се увеличава (например при продължително лечение с β-адреноблокер пропронолол, чувствителността на β-адренергичните рецептори към β-адреностимулаторите се увеличава).
Нарушения поради нарушена функция на рецепторите
В допълнение към индивидуалните различия в чувствителността към лекарства, има заболявания, причинени от дисфункция на някои компоненти на механизма на вътреклетъчно предаване на сигнал от рецептора към ефектора. Със загубата на функция на високоспециализирани рецептори фенотипните прояви на заболяването могат да бъдат ограничени (например с феминизация на тестисите, свързана с генетично отсъствие или структурни дефекти на андрогенните рецептори). Ако се наруши по-универсален механизъм вътре в предаването на клетъчния сигнал, симптомите на заболяването са по-разнообразни, като например при миастения гравис и някои форми на инсулинорезистентния захарен диабет, причинени съответно от автоимунни дисфункции на N-холинергичните рецептори и инсулиновите рецептори. Дефектите във всеки компонент, участващ в преобразуването на сигнала от много рецептори, водят до множество ендокринни нарушения. Пример е хетерозиготна форма на дефицит на G s протеин, която активира аденилатциклазата във всички клетки (Spiegel и Weinstein, 1995). Хомозиготна форма на дефицит на този протеин може да доведе до смърт.
Нарушенията в структурата или локализацията на рецепторите могат да се проявят като отслабена или засилена реакция към лекарството, както и други нежелани ефекти.
Мутациите, кодиращи генни рецептори, са способни да променят както отговора на еднократна употреба на лекарството, така и ефективността на дългосрочното лечение. Например, дефект на β-адренергичните рецептори, отговорни за отпускането на гладката мускулатура на бронхите и регулирането на резистентността на дихателните пътища, засилва намаляването на чувствителността на тези рецептори към β-адреностимуланти по време на дългосрочно лечение на пациенти с AD. Тъй като мутациите, отговорни за нарушената рецепторна функция, са идентифицирани и съответните гени са клонирани, ще бъде възможно да се разработят методи за лечение на такива заболявания.
Класификация на рецепторите
Традиционно лекарствените рецептори са идентифицирани и класифицирани въз основа на ефектите и относителната активност на селективни агонисти (стимуланти) и антагонисти (блокери), действащи върху тези рецептори. Например, ефектите на ацетилхолин, които се възпроизвеждат при взаимодействие с холинергичните рецептори на мускариновия алкалоид и са блокирани от атропин, се наричат \u200b\u200bмускаринови ефекти, а ефектите, които се възпроизвеждат при взаимодействие с холинергичните рецептори на никотина, се наричат \u200b\u200bникотинови ефекти. Рецепторите, които медиират ефектите на мускарин и никотин, се наричат \u200b\u200bсъответно М и N холинергични рецептори. Въпреки че такава класификация обикновено не отразява механизма на действие на лекарствата, е удобно да се систематизира тяхното действие. Всъщност твърдението, че лекарството стимулира рецептори от определен тип, едновременно определя спектъра на ефектите на дадено лекарство и вещества, които усилват или отслабват тези ефекти. Валидността на такива претенции обаче може да се промени с идентифицирането на нови видове и подтипове рецептори, откриването на допълнителни механизми на действие на лекарства или неизвестни досега странични ефекти.
Рецепторни подтипове
С появата на все по-голямо разнообразие от силно селективни лекарства стана ясно, че известните досега видове рецептори са разделени на много подтипове. Методите за молекулно клониране се превърнаха в значителна помощ при изучаването на нови подтипове на рецептори, а получаването на рекомбинантни рецептори улесни създаването на лекарства, които селективно действат на тези рецептори. Различните, но свързани с тях подтипове рецептори често (макар и не винаги) взаимодействат с различни агонисти и антагонисти. Рецепторите, за които не са идентифицирани селективни агонисти или антагонисти, обикновено не принадлежат към един подтип, а към изоформи от същия рецептор. Отделните подтипове могат също да се различават в механизмите на вътреклетъчно предаване на сигнал. М1 и МЗ холинергичните рецептори например действат чрез протеина G q, който активира фосфолипаза С, косвено причинява освобождаването на Са 2+ от вътреклетъчните депа и М2 и М 4 холинергичните рецептори чрез протеина G i, който инхибира аденилатциклазата. В същото време разделянето на рецепторите на видове и подтипове често се определя не от механизма на действие, а от случаен избор или се основава на установени идеи. И така, α 1 -, α 2 - и β-адренергичните рецептори се различават по реакция на лекарства и при предаване на сигнал (активират съответно протеини G i, G q и G s), въпреки че α и β-адренергичните рецептори са от различен тип, т.е. и α 1 - и α 2 -адренорецептори - към различни подтипове в рамките на един и същи тип. Изоформите на α 1 -адренорецепторите α 1A, α 1B и α 1D се различават малко по своите биохимични свойства; същото е характерно за подтиповете изоформаза на β-адренергичните рецептори (β 1, β 2 и β 3).
Разликите между рецепторните подтипове се използват за създаване на силно селективни лекарства, например лекарства, които имат различни ефекти върху една и съща тъкан поради свързване с рецепторни подтипове, които се различават по механизмите на вътреклетъчно предаване на сигнала. В допълнение, лекарствата могат селективно да са насочени към определени клетки или тъкани, експресиращи рецептори на подтип. Колкото по-голяма е селективността на лекарствата (във връзка с определена тъкан или във връзка с определен ефект), толкова по-благоприятно е съотношението на нейните ползи и нежелани ефекти.
С помощта на молекулярно-генетични методи са открити не само различни изоформи на рецептори, но и гени, кодиращи нови, неизвестни досега рецептори. Много от тези рецептори вече са причислени към един или друг известен клас и тяхната функция е изследвана с помощта на съответните лиганди. Все още не са намерени лиганди за някои рецептори.
Откриването на много изоформи на един и същ рецептор, кодирани от различни гени (особено ако изоформите не се различават по механизмите на вътреклетъчно предаване на сигнал и взаимодействат със същите ендогенни лиганди) позволява експресията на рецептори в различни клетки да бъде независимо регулирана в съответствие с нуждите на организма в различни възрастови периоди.
Нерецепторно медиирано действие на лекарството
Не всички лекарства действат чрез макромолекулните структури - рецептори. Някои лекарства взаимодействат с малки молекули или йони, които присъстват в организма нормално или в едно или друго патологично състояние. И така, антиацидите неутрализират солната киселина в стомаха. Месна (лекарство, което бързо се отделя от бъбреците и неутрализира свободните радикали) се свързва с активните метаболити на някои противоракови лекарства, намалявайки тежестта на нежеланите реакции от пикочните пътища. Редица биологично неактивни вещества (например манитол) могат да бъдат въведени в количества, достатъчни за увеличаване на осмоларността на биологичните течности и по този начин да променят разпределението на извънклетъчните и вътреклетъчните течности. С помощта на тези вещества е възможно да се увеличи диурезата, да се увеличи bcc, да се елиминира мозъчният оток. В допълнение, те се използват като слабителни.
Някои лекарства могат да се интегрират в компонентите на клетката и да променят функциите си поради структурни сходства с веществата, които съставляват тези компоненти. Например, аналози на пурини и пиримидини се вкарват в нуклеинови киселини и се използват като антивирусни и противотуморни средства.
AP Викторов "Клинична фармакология"
По правило механизмът на действие на лекарствата се основава на тяхната способност да инициират (задействат) сложни биохимични n / или биофизични процеси, които в крайна сметка променят и / или оптимизират функционалната активност на целевата клетка.
Лекарствата могат да осъществят действието си срещу органи и / или прицелни клетки чрез:
Пряко химическо взаимодействие;
Физико-химично взаимодействие върху клетъчната мембрана;
Действия върху специализирани ензими;
Действия върху регулаторните гени;
Действия върху специфични рецептори.
Пряко химическо взаимодействие LS. Този механизъм на действие на лекарствата е доста рядък и може да се реализира извън клетката, например в лумена на стомаха или червата. Същността му се състои във факта, че лекарствата влизат в директна химическа реакция с молекули и / или йони, които се образуват в организма в нормално състояние, когато възникне патологично състояние. Пример за пряко химическо взаимодействие е химическата реакция на неутрализиране на солна киселина на стомаха при приемане на антиацидни лекарства (вж. Т. 2, стр. 112).
Физико-химично взаимодействие на лекарства върху клетъчната мембрана. Една от основните функции на цитоплазмената мембрана е осъществяването на йонен обмен между цитоплазмата и извънклетъчната среда. Трансмембранният йонен обмен може да се извърши и чрез специални трансмембранни йонни канали, зависими от напрежението - натрий, калий, калций, хлор и др. Някои лекарства, достигайки клетъчната мембрана, взаимодействат с тези канали и променят функционалната им дейност. Така например, антиаритмичният ефект на лекарство от клас IA, хинидин, се основава на способността му да блокира преминаването на Na + йони през трансмембранни натриеви канали (виж Т. 2, стр. 35).
Ефектът на лекарствата върху специализираните ензими. Сравнително малко количество лекарства реализира фармакологичния му ефект чрез промяна на активността на някои специализирани клетъчни ензими. Лекарствата, които повишават активността на клетъчните ензими, се наричат \u200b\u200bензимни индуктори. Такова действие се притежава например от хапчета за сън и антиконвулсантно лекарство фенобарбитал, което значително засилва активността на микрозомалните чернодробни ензими. Биологичното значение на този ефект на фенобарбитала и близки до него LS ще бъде разгледано по-долу.
Лекарствата, които инхибират активността на специализирани ензими, се наричат \u200b\u200bензимни инхибитори. Така например, антидепресант от групата на инхибиторите на моноаминооксидазата (МАО), лекарството пирлиндол реализира антидепресантния си ефект чрез потискане на активността на ензима МАО в централната нервна система (виж Т. 1, стр. 294).
Способността да инхибира активността на ензима ацетилхолинестераза е в основата на фармакологичната активност на антихолинестеразните лекарства, например физостигмин. Известно е, че при физиологични условия ацетилхолинестеразата инактивира (унищожава) ацетилхолин, невротрансмитер, който предава възбуждане в синапсите на парасимпатиковата нервна система. Физостигминът, потискащ активността на ацетилхолинестеразата, насърчава натрупването в синапсите на парасимпатиковата система на невротрансмитера ацетилхолин, в резултат на което се повишава тонусът на парасимпатиковата нервна система, което се проявява на системно ниво чрез развитието на брадикардия, понижаване на кръвното налягане (на гастроинтетичния тракт), на стомашно-чревния тракт (понижаващо кръвно налягане) ученик и т.н.
Лекарствата могат да взаимодействат обратимо и необратимо с ензимите. Например, лекарството еналаприл обратимо инхибира активността на ангиотензин конвертиращия ензим, което в частност води до понижаване на кръвното налягане, докато токсичните вещества от органофосфор необратимо инхибират активността на ацетилхолинестеразата.
Ефектът на лекарствата върху регулаторните гени. В момента учените правят опити да създадат лекарства, които реализират фармакологичните им ефекти, като влияят пряко върху физиологичната активност на регулаторните гени. Тази тенденция изглежда особено обещаваща, след като структурата на човешкия геном беше дешифрирана през 2000 г. Смята се, че селективното нормализиране на функцията на регулаторните гени под въздействието на лекарства ще направи възможно постигането на успех в лечението на много, включително нелечими преди това заболявания.
Ефектът на лекарствата върху рецепторите. Преди да преминем към спецификата на взаимодействието на лекарствата с рецепторите, е необходимо да се изясни какво разбираме под термина „рецептор“ (от латински recipio - вземете, вземете).
От курса на физиологията е известно, че терминът "рецептор" означава високоспециализирани формации, които са в състояние да възприемат, трансформират и предават енергията на външен сигнал към нервната система. Такива рецептори се наричат \u200b\u200bсензорни (от лат. Sensus - усещане, усещане, възприятие).
Сензорните рецептори включват рецептори на органите на слуха, зрението, обонянието, вкуса, допира и т.н. Сензорните рецептори на тези органи принадлежат към така наречените екстерорецептори.
Ако наличието на сетивни органи, които реагират на външни стимули на дразнене, е било известно от древни времена, то присъствието на сензорни рецептори вътре в тялото е било под въпрос до средата на 19 век. За първи път присъствието на такива рецептори вътре в тялото беше предложено от руския физиолог I.F.Pion, който през 1866 г. показа спад на кръвното налягане поради дразнене на аортата при експеримент със заек. Това откритие породи търсенето и изучаването на рецептори, разположени вътре в тялото, и самите тези рецептори бяха наречени интерорецептори.
До началото на 20 век беше разкрит достатъчен брой сензорни интерорецептори и е доказана тяхната важна роля в регулацията на физиологичните функции на организма.
През 1905 г. J. Langley доказва, че когато лекарството се прилага върху клетъчната мембрана, се развива фармакологичен ефект, ако се прилага само върху конкретна област от него. Освен това този сайт представлява само малка част от общата площ на клетъчната повърхност. Това наблюдение позволи на J. Langley да заключи, че на клетъчната мембрана съществуват специализирани рецепторни сайтове, взаимодействащи с лекарства.
Въпреки това приоритетът при създаването на рецепторната теория за действието на лекарствата принадлежи на немския физиолог П. Ерлих, който през 1906 г. въвежда понятието „рецептор“ и формулира постулата „лекарството не работи, ако не се фиксира върху клетъчната мембрана“. Според теорията на P. Ehrlich, молекулата на лекарството има две функционално активни групи, едната от които осигурява фиксацията му върху клетъчната повърхност в областта на лекарствения рецептор, а втората функционална група взаимодейства с рецептора и задейства сложна верига от биохимични реакции, които променят неговата (клетъчна) физиологична активност ,
По този начин още в началото на 20 век. стана ясно, че има поне два класа интерорецептори: сензорни рецептори, които предават информация за състоянието на вътрешните органи и телесните тъкани към централната нервна система; маркиращи рецептори, които взаимодействат с лекарства, които променят функционалната активност на целевите клетки.
Веднага трябва да се отбележи, че в бъдеще в текста на учебника, за да се избегне объркване в терминологията, рецепторите за лекарства и биологично активни вещества, т.е. белязани или циторецептори. ще бъдат обозначени с термина "рецептор", докато сензорните интерорецептори ще бъдат обозначени с термин, характеризиращ тяхната функционална активност, например "барорецептори", "рецептори на болката" и т.н.
Откритието от П. Ерлих върху клетъчната мембрана на лекарствените рецептори послужи като отправна точка за развитието на фармакологичната наука, по-специално фармакодинамиката, една от основните задачи на която е да се проучат рецепторните механизми на действие на лекарствата.
Понастоящем са разкрити структурата на голям брой клетъчни рецептори, особеностите на взаимодействието на определени биологично активни съединения с тях, което даде възможност, от една страна, да се разбере механизмът на действие на известни лекарства, а от друга, беше основата за създаването на нови високоефективни лекарства.
Естествено е трудно да си представим, че в хода на еволюцията в човешкото тяло се образуват рецептори за различни синтетични (химически получени) лекарства, особено след като огромното мнозинство лекарства, представени на съвременния фармацевтичен пазар, са синтезирани през последните 50 години или по-малко. Доказано е, че рецепторният апарат на клетката е много древна функционално-структурна формация. И така, a- и β-адренорецепторите (рецептори, взаимодействието на които норепинефрин и адреналин влияят върху функционалната активност на клетката) се намират не само в клетките на животните, но и в клетъчните мембрани на растителните клетки, например в клетките на растителната нителла, където a- и β- адренореценторите регулират движението на протоплазма (клетъчното съдържание).
Тогава какви са рецепторите за лекарства, открити от П. Ерлих, и защо те взаимодействат с тях?
В момента няма съмнение, че така наречените лекарствени рецептори всъщност са рецептори за ендогенни (произвеждани в организма) биологично активни вещества, участващи в регулирането на функционалната дейност на вътрешните органи и телесните тъкани. Такива биологично активни съединения включват вещества, освободени от нервните окончания в момента на предаване на нервния сигнал, както и хормони, витамини, аминокиселини и др. За всяко ендогенно биологично активно вещество има строго специфични рецептори за него. Така, например, биологично активното вещество, произведено в организма, адреналинът, може да активира строго специфични a- и β-адренорецептори, а глюкокортикостероидите - хормони на надбъбречната кора - взаимодействат само с стриктно специфични за тях глюкокортикостероидни рецептори.
Синтетичните лекарства, които реализират ефектите си, взаимодействайки с рецепторния апарат на клетката, по своята химическа структура са повече или по-малко подобни на ендогенни биологично активни съединения, които взаимодействат с подобни рецептори. Например синтетичните вазоконстрикторни (причиняващи вазоконстрикция) лекарства фенилефрин са близки по своята химическа структура с ендогенното биологично активно вещество норепинефрин, следователно, подобно на норепинефрин, той има способността да стимулира a-адренорецепторите.
Понякога, поради особеностите на тяхната химическа структура, лекарствата могат да взаимодействат не със самия рецептор, а със съседната част на клетъчната мембрана. Тъй като в този случай лекарството не взаимодейства със самия рецептор, а със съседната част на клетъчната мембрана, те говорят не за възбуждащ или блокиращ ефект върху рецептора, а за алостеричен (от гръцки alios - друг, различен) ефект или ефект. В резултат на това може да настъпи промяна както в структурата на мембраната, съседна на рецептора, така и в отделните компоненти на самия рецептор, което може да доведе до промяна в чувствителността на рецептора към биологично активно вещество, специфично за него. В случаите, когато чувствителността на рецептора към биологично активно вещество се увеличава, те говорят за сенсибилизация (от латински sensus - смисъл) или сенсибилизация (от латински sensibilis - чувствителност) на рецептора, а в случаите, когато чувствителността на рецептора намалява, те говорят за десенсибилизация рецептор.
Особеността на алостеричния ефект се крие във факта, че лекарствата, които имат този вид механизъм на действие, не влияят пряко върху предаването на нервен импулс, а го променят в желаната посока. Например механизмът на действие на анксиолитиците (лекарства против тревожност; синоним: транквилизатори), които по своята химическа структура са производни на бензодиазепин, се основава на феномена на алостеричното възбуждане на постсинаптичните бензодиазепинови рецептори. Възбуждането на последните от своя страна насърчава активирането на инхибиторните постсинаптични рецептори на гама-аминомаслена киселина (GABA рецептори), което се проявява клинично чрез елиминиране на симптоми на невротични заболявания като тревожност, тревожност, страх и др.
Рецепторите, взаимодействащи с които биологично активно вещество или лекарство по някакъв начин променя функционалното състояние на целева клетка, се наричат \u200b\u200bспецифични.
В допълнение към специфичните рецептори се изолират така наречените лекарствено-специфични рецептори. В специализираната медицинска литература тези рецептори се наричат \u200b\u200bоще „мястото на загубата“ на лекарствата. Свързвайки се с такива рецептори, лекарствата нямат никакъв биологичен ефект, но самите те стават биологично неактивни. Пример за този тип рецептори могат да служат като рецептори, разположени върху плазмените протеини, по-специално върху водоразтворимите протеини - албумин. Значението на това явление ще бъде разгледано подробно по-долу (вж. Т. 1, стр. 72).
Структурата на рецепторите е доста сложна, но повечето от тях са протеинови макромолекули или гликопротеини, които също могат да включват йони, липиди, нуклеинови киселини и др. Рецептор, т.е. образуващата го протеинова макромолекула се характеризира със специфично, специфично за всеки рецептор, пространствено разположение на неговите химични групи. Протеиновата макромолекула, образуваща рецептора, може да бъде интегрирана (потопена) в липидната двуслойна на цитоплазмената мембрана или локализирана вътре в клетката. Основната функция на клетъчния рецептор е да „разпознава“ химичен сигнал, предаван към него чрез ендогенно биологично активно вещество и / или лекарства и да го трансформира в съответния биохимичен и / или биофизичен отговор на клетката.
По-рано се смяташе, че лекарствата или ендогенните биологично активни вещества взаимодействат с рецепторите от типа „ключ и заключване“, т.е. рецепторът има такава структура, която позволява на лекарството да намери "вашия" рецептор, да се свърже с него и, както би било, "включете" и "изключете". С развитието на медицинската наука обаче стана ясно, че егото не е съвсем така. Понастоящем молекулните процеси на превръщане на извънклетъчните сигнали във вътреклетъчна, регулираща клетъчната функция, вече са проучени доста добре. механизми, които водят до ефекта от взаимодействието на ендогенни биологично активни вещества или лекарства с рецептори.
При взаимодействие с рецептора на ендогенно биологично активно вещество и / или активно L C като него, възниква конформация - пространствена промяна под формата на протеинова макромолекула, която е спусъкът на различни вътреклетъчни процеси, които определят реакцията на целева клетка към медиатор и / или лекарство. Например, активирането на адренергичните рецептори на гладката мускулатура на бронхите под въздействието на β2-адреностимулаторния фенотерол води до повишаване на активността на ензима аденилатциклаза, което допринася за натрупването на цикличен аденозинов монофосфат (сАМФ) в клетката и в резултат на това клетъчна релаксация.
Като цяло биологичните термини, клетъчните рецептори могат да се считат за строго специализирани „сетивни органи“ на клетките, чрез които те възприемат „информация“, излъчвана например от централната нервна система и / или ендокринната система. Въпреки важната роля на рецепторния апарат, рецепторите заемат само незначителна част от клетъчната мембрана. Например, М-холинергичната рецепторна клетка заема не повече от 1/6 000 от нейната повърхност.
Изучаването на характеристиките на взаимодействието на лекарствата с рецептора, от една страна, ни позволява да разберем основата на молекулярния механизъм на неговото действие, а от друга, дава информация какви промени трябва да се направят в структурата на лекарствата, за да се повиши способността му да взаимодейства с този рецептор, т.е. , позволява целенасочен синтез на нови високоефективни лекарства.
При физиологични условия различните клетъчни рецептори не функционират независимо, но са в постоянно взаимодействие помежду си, като по този начин регулират специфичната активност на клетката. Например, активирането на сърдечните β-адренорецептори от ендогенни норепинефрин причинява по-специално увеличаване на броя на сърдечните контракции и активирането на сърдечните М-холинергични рецептори от ендогенния ацетилхолин, напротив, причинява намаляване на броя на сърдечните контракции.
Голям принос за разбирането на рецепторните механизми на действие на лекарствата направиха откриването на пре- и постсинаптични рецептори. Синапс (от гръцки synapsis - връзка, връзка) е специализирана контактна зона между нервните клетки или други възбудими структури на тялото, която осигурява предаването на входяща информация и запазването на нейното информационно значение. Изучаването на структурата и функционалната роля на синапсите започва в края на 19 век. след това испанският хистолог S. Ramon n Cajal (S. Ramon at Cajal) предложи наличието на специализирана система за предаване в централната нервна система. Синапсите получиха името си през 1897 г., когато английският физиолог К. Шерингтън предложи този термин да се отнася до зоната на контакт между нервните клетки.
В момента има три типа синапси:
1) „електрически“ синапси, при които информацията се предава чрез прехвърляне на електрически сигнал от пресинаптична мембрана. Този тип синапс се нарича efaps (от гръцки. Ephapsis - стегнат контакт);
2) „химични“ синапси, при които информацията се предава чрез специални биологично активни вещества - невротрансмитери (от гръцки. Neuron - нерв и латински. Mediator - медиатор; синоним: медиатор);
3) „смесени“ синапси, при които информацията се предава както химически, така и електрически.
Фармакологичните ефекти на огромното мнозинство лекарства, които влияят върху функциите на синапсите, се реализират чрез ефекта им върху гот или друг етап на предаване на сигнала в химичните синапси, т.е. в синапси от втория вид.
По правило химичните синапси се класифицират от невротрансмитери, които предават нервни импулси, както следва:
Синапсите, в които ацетилхолинът действа като медиатор, се наричат \u200b\u200bхолинергични;
Синапси, при които адреналинът и норепинефринът действат като медиатор, се наричат \u200b\u200bадренергични;
Синапсите, при които АТФ и аденозинът действат като медиатор, се наричат \u200b\u200bпуринергични;
Синапсите, в които гама-аминомаслената киселина действа като медиатор, се наричат \u200b\u200bGABA-ергични и т.н.
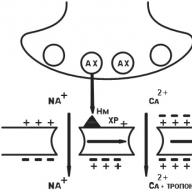
В момента структурата на синапса е добре разбрана. Синапсът се състои от пресинаптичен процес на нервна клетка (аксонов край) и "сигнал" -приемащ апарат, разположен върху мембраната на ефекторна ("изпълнителна") клетка.
Аксонът на еферентния неврон, приближаващ се до ефекторната клетка, губи миелиновата обвивка, разширява се и образува така нареченото пресинаптично удебеляване (фиг. 1.5). Повърхността на нерва, завършваща към клетъчната мембрана на ефекторната клетка, се нарича пресинаптична мембрана. Мястото на ефекторната клетка срещу пресинаптичната мембрана се нарича постсинаптична мембрана (виж фиг. 1.5). В зависимост от структурните особености на синапса пресинаптичната мембрана може да има повече или по-малко гънки и по този начин да има по-голяма или по-малка площ. В химичните синапси пресинаптичната мембрана не контактува директно с постсинаптичната мембрана, а се отделя от нея на малко разстояние, наречено синаптична цепнатина (виж фиг. 1.5).
Пресинаптично сгъстяване, т.е. крайната част на аксона съдържа по-голям брой митохондрии, вътреклетъчни органели, участващи в синтеза и натрупването на енергия, която е по-голяма от тялото на неврона, което показва интензивността на енергийните процеси, протичащи в този участък на нервната клетка. В допълнение към митохондриите, пресинаптичните удебелявания съдържат голям брой малки везикули - везикули. Средно около 20 000 везикули се съдържат в едно пресинаптично удебеляване. Последните са локализирани в пресинаптичното сгъстяване неравномерно, като правило повечето от тях са разположени близо до пресинаптичната мембрана. Невротрансмитерът се синтезира в тялото и аксона на неврона и се натрупва във везикулите. Всеки везикул съдържа няколко хиляди молекули на невротрансмитер (от I 000 до 50 000). Когато възникне нервен импулс, везикулата се слива с пресинаптичната мембрана и невротрансмитерът се секретира в синаптичната цепнатина (вж. Фиг. 1.5).
Фиг. 1.5. Принципна схема на структурата на "химичния" синапс:
a е схематично изображение; б - електронна микрография; 1- пресинаптичен нервен завършек; 2 - prssynaptic мембрана; 3 - постсинаптична мембрана; 4 - синаптична цепка; В - везикул; NM - невротрансмитер; P - постсинаптичен рецептор: OZ - „обратно“ улавяне на невротрансмитер; SF е специализиран ензим, който унищожава излишния невротрансмитер в синаптичната цепнатина
Функционално активните рецепторни образувания са разположени върху постсинаптичната мембрана, които са в състояние да взаимодействат с невротрансмитера, освободен от пресинаптичната мембрана по време на преминаването на нервен импулс. Рецепторите, разположени на постсинаптичната мембрана, се наричат \u200b\u200bсинаптични или постсинаптични рецептори в специализирана медицинска литература. Под постсинаптични рецептори се разбира макромолекули с протеинов характер, вградени в постсинаптичната мембрана с генетично предопределена структура и функция, способни да обратимо взаимодействат с невротрансмитери и / или лекарства поради функционалните групи на активния център („разпознаващата“ част от макромолекулата).
Предаването на нервния сигнал в синапса става по следния начин: под въздействието на нервен стимул везикулите се придвижват към пресинаптичната мембрана и невротрансмитерът се секретира от екзоцитозата в синаптичната цепнатина (вж. Фиг. 1.5). Невротрансмитерът, освободен в синаптичната цепнатина, достига до постсинаптичната мембрана, където, взаимодействайки с постсинаптичния рецептор, той задейства верига от биохимични и / или биофизични реакции, резултатът от които е физиологичният отговор на целевата клетка. Въпреки това, не цялото количество освободен невротрансмитер достига постсинаптични рецептори и взаимодейства с тях. Част от невротрансмитера се улавя от пресинаптичната мембрана и се „връща“ на местата за съхранение. Това явление се нарича феномен на обратното захващане на невротрансмитерите.
Останалото количество не взаимодействащ невротрансмитер рецептор се унищожава в синаптичната цепнатина от специализирани ензими. Това явление се нарича разграждане на невротрансмитерите. Например ензимът ацетилхолинестераза катализира (ускорява) процеса на разграждане (разрушаване) в синаптичната цепнатина на невротрансмитера ацетилхолин.
За разлика от невротрансмитера, неговите метаболитни продукти имат невротрансмитер активност. Целият процес на взаимодействие на невротрансмитера с рецепторите и разрушаването на неговия излишък от специфичен ензим е изключително кратък и не надвишава 2 ms (1 ms \u003d 0,001 s).
Такава кратка продължителност на този процес се обяснява, от една страна, с изключително бързото освобождаване на невротрансмитера от рецептора, а от друга, с високата скорост на ензимно инактивиране на невротрансмитера в синаптичния шел.
Основно функционалната дейност на синапса може да бъде променена, както следва:
За ускоряване, намаляване или блокиране на синтеза, натрупването и / или катаболизма (разрушаването) на невротрансмитера в пресинаптичния завършек. В резултат на това съдържанието на невротрансмитера и вследствие на това интензивността на неговата физиологична активност по някакъв начин ще се промени.
Например, симпатолитичният резерпин предотвратява натрупването на катехоламини в синаптичните везикули до пълното им изпразване. В резултат на това количеството на невротрансмитер норепинефрин, освободен в синаптичната цепка, рязко спада. На системно ниво този ефект се реализира под формата на понижение на кръвното налягане. Някои лекарства не влияят пряко върху съдържанието на невротрансмитери в пресинаптичния завършек, но инхибират активността на ензимите, които ги унищожават. Така действат редица антидепресанти. Например, антидепресантът пирлиндол инхибира (потиска) активността на ензима моноамин оксидаза при пресинаптичното прекратяване и в резултат на това повишава концентрацията на невротрансмитери като норепинефрин, допамин и серотонин в него. Клинично този ефект на пирлиндол се проявява чрез намаляване на чувствата на тревожност и страх, подобрено настроение, повишена физическа активност и др .;
Променете (улеснете, усложете) способността на невротрансмитера да проникне през пресинаптичната мембрана и следователно увеличете или намалете количеството на невротрансмитера, освободено в синаптичната цепнатина с всеки импулс.
Например, психостимулиращият амфетамин улеснява освобождаването на катехоламините в адренергичните синапси на централната нервна система и по този начин увеличава съдържанието им в синаптичната цепнатина. Клинично този ефект на лекарството се проявява в подобрено настроение, усещане за прилив на сила, повишена ефективност. Тетаничният токсин блокира отделянето на инхибиторни невротрансмитери (GABA, глицин) в централната нервна система и по този начин рязко намалява съдържанието им в синаптичната цепнатина, което се проявява клинично с развитието на припадъци;
Блокирайте или стимулирайте обратното поемане на невротрансмитерите от пресинаптичната мембрана и следователно увеличете или намалете концентрацията на невротрансмитери в синаптичната цепнатина.
Например трицикличният антидепресант имипрамин блокира обратното захващане на невротрансмитера норепинефрин от пресинаптичната мембрана и по този начин рязко повишава концентрацията му в синаптичната цепнатина. Клинично този ефект на имипрамин се проявява чрез подобрено настроение, повишена умствена и физическа активност;
Стимулирайте или блокирайте активността на ензимите, които унищожават невротрансмитера в синаптичната цепнатина.
Например, антихолинестеразното лекарство физостигмин намалява активността на ензима ацетилхолинестераза, който унищожава невротрансмитер ацетилхолин в синаптичната цепка и по този начин допринася за повишаване на концентрацията му, което може да се прояви клинично, по-специално чрез намаляване на вътреочното налягане и свиване на зеницата.
Стимулират или блокират постсинаптичните рецептори, т.е. имитира или блокира ефекта на невротрансмитерите.
Например наркотичните аналгетици, които възбуждат постсинаптичните опиоидни рецептори и по този начин имитират влиянието на невротрансмитерите - енкефалините. Стрихнинът, като блокира рецепторите на инхибиторния невротрансмитер глицин, възпрепятства реализирането на неговия инхибиторен ефект, в резултат на което стрихнин във високи дози предизвиква гърчове.

Фиг. 1.6. Схематично представяне на локализацията на пре- и постсинаптичните рецептори, както е показано от адренергичния синапс (обяснение в
NM - невротрансмитер; М2 (-) - холинергичен "инхибиторен" пресинаптичен хетерорецептор; β 1 (+) - адренергичен „активиращ“ пресинаптичен авторецептор; β - адренергичен постсинаптичен рецептор
В допълнение към рецепторите, разположени на постсинаптичната мембрана, т.е. постсинаптични рецептори, рецептори, разположени на пресинаптичната мембрана, т.е. пресинатични рецептори (фиг. 1.6). Въпреки факта, че както пре-, така и постсинаптичните рецептори могат да бъдат възбудени от един и същ невротрансмитер, функционалната роля на тези рецепторни образувания в синапсите е различна. Ако постсинаптичните рецептори са крайната връзка за предаване на нервен импулс към ефекторния орган, т.е. осигуряват еднопосочна проводимост на нервен импулс от центъра към периферията, тогава пресинаптичните рецептори участват в
регулиране на невротрансмитерната активност на синапса, т.е. до известна степен влияят върху процесите на освобождаване и / или синтез на невротрансмитер в него. Трябва да се подчертае, че пресинаптичните рецептори не участват пряко в провеждането на нервен импулс от неврон към ефекторния орган.
Пресинаптичните рецептори се разделят на две големи групи: авто- и хетероневромодулиращи рецептори (вж. Фиг. 1.6).
Пресинаптичните авторецептори включват рецептори, които се възбуждат от собствения си невротрансмитер за този синапс.
Например в синапси, локализирани в зоната на контакт между соматичните нерви и набраздения мускул, когато в синаптичната цепнатина има излишък от ацетилхолин, взаимодействайки с пресинаптични авторецептори, той инхибира освобождаването на нова част от невротрансмитера възбуждането на пресинаптичните авторецептори регулира освобождаването на ацетилхолин от пресинаптичните терминали.
Но върху пресинаптичната мембрана, в допълнение към авторецепторите, т.е. рецептори, които са чувствителни към невротрансмитер, който предава възбуждане в даден синапс, могат да бъдат разположени рецептори, които не са чувствителни към невротрансмитер, който предава възбуждане в даден синапс, но които взаимодействат с друг тип невротрансмитер.
Например върху синаптичната пресинаптична мембрана, в която ацетилхолинът е невротрансмитерът, могат да бъдат разположени пресинаптични рецептори, чувствителни към невротрансмитера норепинефрин. Този тип пресинаптичен рецептор се нарича хетероневромодулиращ рецептор.
По този начин синапсът представлява сложна анатомична и функционална формация, която осигурява предаването на нервен импулс от неврон към неврон или от неврон към ефекторна клетка.
Последователността на функционалната активност на синапса (етапи на синаптично предаване) е следната:
Синтез и натрупване на невротрансмитер във везикули, локализирани в пресинаптични сгъстявания (синтезът на невротрансмитер се случва не само в пресинаптичните сгъстявания, но и в невроните и аксоните);
Освобождаването на невротрансмитера в синаптичната цепнатина в момента на преминаване на нервен импулс;
Взаимодействието на невротрансмитера с постсинаптичните рецептори, което води до активиране на рецепторите и промяна във функционалната активност на ефекторната клетка;
Инактивирането на невротрансмитера (ензимно) и / или обратното му поемане от пресинаптичната мембрана, т.е. възстановяване на способността на синапса отново да предава нервен импулс към ефекторна клетка.
Синапсите имат следните основни свойства:
Едностранно провеждане на възбуждане (нервен импулс може да премине само от пресинаптичната мембрана към постсинаптичната);
Синаптично забавяне, т.е. определено време се изразходва за предаване на нервен импулс в синапса. (Скоростта на синаптичното предаване е средно повече от 10 пъти по-ниска от скоростта на разпространение на нервен импулс през нерва. За химичен синапс обикновено е от 0,2 до 0,5 ms);
Умора - постепенно намаляване или пълно спиране на предаването на нервен импулс с продължителна нервна стимулация. Основата на това явление е, от една страна, изчерпването на резервите на невротрансмитерите при пресинаптични сгъстявания, а от друга страна, намаляването на чувствителността към постсинаптичните рецептори към невротрансмитера;
Висока чувствителност на синаптични образувания към лекарства и отрови.
Именно върху последното свойство на синапсите се основава цялата фармакология на лекарствата, които влияят върху функционалната активност на синапсите, разположени в различни органи и тъкани на тялото. Трябва да се подчертае, че обектът на фармакологичното действие може да бъде всеки от етапите на синаптично предаване. Като лекарства, които влияят на синаптичното предаване, се използват екзогенни аналози на невротрансмитерите, техните химически прекурсори и други биологично активни вещества, които по всякакъв начин могат да променят функционалната активност на синапса.
Трябва да се отбележи, че много лекарства имат не една, а няколко точки на приложение на ефекта на нивото на синапса. Така например, антидепресантът пирлиндол не само инхибира активността на ензима моноамин оксидаза в синаптичната цепка, но и блокира обратното поемане на норепинефрин от пресинаптичната мембрана.
Във връзка с локализацията на рецептора към синапса те могат да бъдат разделени на пресинаптични, постсинаптични и екстрасинаптични. Последните например включват рецептори, разположени върху клетъчните мембрани на тромбоцитите.
От гледна точка на клетъчната топография (местоположение), рецепторите могат също да бъдат класифицирани според тяхното местоположение върху клетъчните структури, както следва:
мембранни рецептори - рецептори, разположени на цитоплазмената мембрана;
цитозолни рецептори - рецептори, разположени върху вътреклетъчни образувания;
ядрени рецептори - рецептори, разположени на мембраната на клетъчното ядро.
Както бе отбелязано по-рано, в резултат на взаимодействие с рецептора на ендогенни биологични вещества или лекарства, функционалната активност на целевите клетки се променя. Този процес може да се осъществи по различни начини, строго определени за различни видове рецептори. В съответствие с това в момента се разграничават четири типа рецептори, всеки от които има свой механизъм, коренно различен от останалите, чрез който сигнал от рецептора инициира каскада от биохимични и / или биофизични реакции, водещи до промяна във функционалното състояние на целевите клетки.
Първите три типа рецептори са локализирани върху клетъчната (цитоплазмена) мембрана, а четвъртият тип рецептори включва цитозолни и ядрени рецептори.
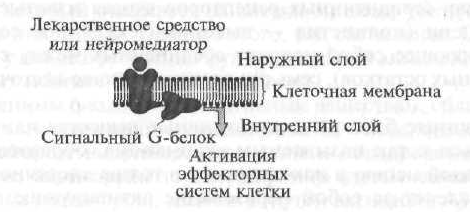
Рецепторите от тип I включват клетъчни (мембранни) рецептори, които реализират ефектите си чрез така наречените сигнални G-протеини (фиг. 1.7).
На първия етап биологично активното вещество или лекарство, „отиващо“ към клетъчната мембрана, „разпознава“ рецептора и взаимодейства с него, след което рецепторът активира специализиран сигнален G протеин, разположен на вътрешната повърхност на мембраната. Освен това активираният G-протеин променя функционалната активност на вътрешния ефектор, който по правило е ензими. Тогава ефекторният елемент, който е ензим, активира вторичен пратеник или вторичен пратеник, който задейства каскада от биохимични реакции, които променят функционалната активност на целевите клетки.
Клетъчни рецептори тип I, т.е. рецепторите, конюгирани с сигнални G-протеини, са структурно сходни един с друг и по своята пространствена организация те са серпентинова (от френски серпантин - змия, топка) структура (фиг. 1.8).
Фиг. 1.7. Структурата на рецептора тип 1 (обяснение в текста)
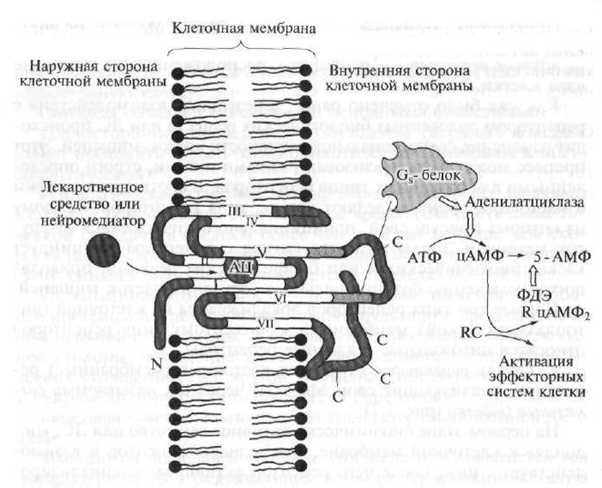
Фиг. 1.8. Схематично представяне на структурата на "серпентина"
рецептор:
N е полипептидната част на рецептора, разположена над клетъчната мембрана; С е полипептидната част на рецептора, разположена под клетъчната мембрана; АС - активният център на рецептора, с който лекарството взаимодейства; отидете невротрансмитер; ATP - аденозин трифосфат - вторичен пратеник; cAMP - цикличен аденозин монофосфат; 5-AMP - аденозин-5 "монофосфат; PDE - фосфодиестераза; R. RC - cAMP-зависим ензим (протеин киназа) с регулаторни и каталитични (ускоряващи реакцията) субединици; 1-VII - полипептидни вериги на серпентин рецептор
Серпентиновите рецептори включват криптирани полипептидни вериги (полипептидът е високомолекулно съединение, което е верига от взаимосвързани аминокиселинни остатъци), които проникват в клетъчната мембрана седем пъти.
Ендогенните биологично активни вещества или лекарства могат да се свържат с така наречения „джоб“, образуван от полипептидната верига и разположен в дебелината на клетъчната мембрана, което води до образуването на активиращ сигнал, който се предава на части от рецепторната верига, разположени в цитоплазмата на клетката. Сигналите G протеини взаимодействат с cy-
стол (вътреклетъчни) секции на пода и пептидна верига; активира и стартира каскада от биохимични реакции, които променят неговата функционална активност по отношение на целевата клетка, т.е. инициира първичен фармакологичен отговор.
В момента са известни няколко типа сигнални G протеини.
Сигнал G, -протеини. Тези сигнални протеини като правило активират ефекторния елемент - ензима аденилатциклаза, който от своя страна стимулира синтеза в клетката (от АТФ) на вторичен пратеник - цикличен аденозин монофосфат (сАМФ). Биологичната роля на cAMP като вторичен пратеник е много важна. Например, увеличаването на съдържанието му в сърдечните клетки води до увеличаване на честотата и силата на сърдечните контракции. В допълнение, увеличаването на концентрацията на сАМФ в различни целеви клетки причинява отпускане на гладката мускулатура на кръвоносните съдове и бронхите, мобилизиране на енергийни резерви (разпад на въглехидрати в черния дроб), потиска способността за агрегация на тромбоцитите, понижава тонуса на миометрия (мускула на матката) и пикочния мехур и др.
Редица невротрансмитери, като адреналин (чрез активиране на β-адренорецептори), допамин (чрез активиране на D 1 -допаминови рецептори), аденозин (чрез активиране на рецептори на аденозин А2) принадлежат към ендогенни биологично активни вещества с способността да активират сигнални G s-протеини, хистамин (чрез активиране на хистамин G2 -рецептороп), серотонин (чрез активиране на серотонин 5-НТ 4 -рецептори), както и редица хормони, например вазопресин (чрез стимулиране на V2-вазопресин рецептори) и др.
Сигнал G i -протеини. За разлика от сигналните G s протеини, активирането на сигнални G i протеини не стимулира, но инхибира активността на ефекторния елемент, ензима аденилат циклаза, което води до намаляване на концентрацията на cAMP в целевите клетки на вторичния месинджър. Намаляването на съдържанието на cAMP в целевите клетки причинява намаляване на сърдечните контракции, повишаване на тонуса на кръвоносните съдове и бронхите, т.е. обратен ефект на увеличаването на съдържанието на cAMP върху целевите клетки. В допълнение, редица сигнални G i протеини участват в регулирането на функционалната активност на трансмембранните йонни Са2+ и К + канали.
Редица невротрансмитери, например, адреналин и норепинефрин (чрез активиране на 2-адренорецептори), допамин (чрез активиране на D2 - допаминови рецептори), аденозин (чрез активиране на А1, принадлежат към ендогенни биологично активни вещества със способността да активират сигнални G i-протеини. аденозинови рецептори), ацетилхолин (чрез активиране на М2 и М4 мускаринови рецептори) и др.
Сигнал G ^ протеини. Тези сигнални протеини допринасят за активирането на друг ефектен елемент на таргетните клетки, ензим фосфорилаза С, който от своя страна стимулира образуването на вторични пратеници диацилглицерол (DAG) и инозитол 1,4,5-трифосфат (ITP) в целевите клетки. Първият от тях (DAG) се свързва с клетъчната мембрана и инициира биохимични реакции, участващи в регулирането на контрактилния статус, растежа и деленето на клетките и секрецията на определени хормони от клетките-мишени. Под въздействието на ензима фосфолипаза А2 DAG може да се метаболизира до арахидонова киселина, която участва в синтеза на биологично активни вещества като ейкозаноиди - простагландини, простациклини, тромбоксани, левкотриени (виж Т. I, стр. 478).
Вторият вторичен пратеник, ITF, не се фиксира върху клетъчната мембрана и се премества във вътреклетъчната среда (цитозол), където инициира освобождаването на Са 2+ йони от клетъчни депа, т.е. насърчава прехода на неактивни йони на Са 2+ в активната форма.
Много изследователи смятат йони на Ca 2+ за третичен вестител или посредник. Това се дължи на факта, че ролята на Ca 2+ йони в регулирането на функционалната активност на клетките е много важна. Са 2+ йони могат да влязат в клетката от външната среда чрез специални трансмембранни йонни канали и / или освободени от клетъчни депа. Основното депо (мястото на натрупване на неактивни йони на Са 2+) в клетката е ендоплазменият, или саркоплазматичният ретикулум (ретикулум саркоплазматичен, синоним: ендоплазмен ретикулум - вътреклетъчна органела, представляваща система от тубули и цистерни, разположени в цитоплазмата, ограничена от частта, осигуряваща мембраната; транспорт на вещества в цитоплазмата). Свободните (активни) Са 2+ йони, идващи от саркоплазмения ретикулум до цитоплазмата, взаимодействат с някои Ca2+ свързващи протеини, най-важният от които е калмодулинът. Комплексният "калмодулин-Са 2+" и / или комплекси от йони на Са 2+ с други калциево-свързващи протеини предизвикват каскада от биохимични реакции в клетката. В резултат на това в зависимост от целевите органи, в които се осъществява този процес, се започва повишаване на контрактилната функция на миокарда и скелетните мускули, повишаване на тонуса на гладките мускули на кръвоносните съдове, бронхите и матката, повишаване на секреторната активност на жлезистата тъкан, стимулиране освобождаването на невротрансмитери от нервните окончания и др. , Доказано е също, че йони на Ca 2+ имат способността да увеличават активността на ензимите, участващи в протеиновия, въглехидратния и мастния метаболизъм.
В допълнение към директната връзка между вторичните пратеници - DAG и ITF и следователно сигнални G q протеини, йони на Ca 2+ при физиологични условия имат доста сложно взаимодействие с cAMP вторичния пратеник, активността на който се регулира от сигнали G s и G i протеини. По този начин беше показано, че свободните Са 2+ йони, влизащи в цитоплазмата на нервна клетка през системата калмодулин-Са 2+, инициират намаляване на съдържанието на iAMP в клетката. В същото време, за да се поддържа отвореното състояние на калциевите йонни канали в клетката, са необходими високи концентрации на cAMP, т.е. намаляването на съдържанието на сАМР, инициирано от комплекса калмодулин-Са 2+, води до спиране на приема на свободни Са2+ йони в цитоплазмата. От друга страна, има доказателства, че cAMP вторичният месинджър засилва абсорбцията на свободни Ca 2+ йони от саркоплазмения ретикулум, т.е. насърчава прехода на Ca 2+ йони от свободната, активна форма към свързаната, неактивна форма.
В резултат на увеличаването на съдържанието на вторични пратеници в целевите клетки - DAG и ITF - тонусът на гладките мускули се увеличава, секрецията на жлезите се увеличава, освобождаването на невротрансмитери от пресинаптични окончания се улеснява, способността за агрегиране на тромбоцитите и др.
Ендогенните биологично активни вещества със способността да активират С q сигнални протеини включват невротрансмитери като норепинефрин (чрез активиране на 1-адренорецептори), ацетилхолин (чрез активиране на мускаринови М1 и МЗ рецептори), серотонин (поради активиране на серотонин 5-НТ2а рецептори), хистамин (поради активиране на хистаминови Н1 рецептори), както и други ендогенни биологично активни вещества, например брадикинин и ангиотензин.
Понастоящем в допълнение към изброения сигнал G-протеини (G s, G |, G q) са идентифицирани други сигнални G-протеини - G s, G i, G q, физиологичната роля на които все още е напълно неясна. Но в същото време има доказателства, че например сигналът C o протеин участва в регулирането на функционалната активност на трансмембранните йонни канали.
Функционалната единица на рецепторите от тип 11 е трансмембранен (проникващ през цялата дебелина на клетъчната мембрана) протеин (ензим). Самият рецептор се състои от два еднакви фрагмента, които се наричат \u200b\u200bмономери. Мономерите са разположени на незначително разстояние един от друг, а самият мономер се състои от две функционално активни субединици - домейни, свързани помежду си от полипептиден сегмент, който пресича липидната двуслойна мембрана (фиг. 1.9). А-субединицата на мономера стърчи над външната повърхност на мембраната и е отговорна за свързването на рецептора с биологично активни вещества, а Р-субединицата се потапя в цитоплазмата на клетката.

Фиг. 1.9. Структурата на рецептора тип II (обяснение в текста): 1 - a-субединица на мономера; 2 - β-субединица на мономера
След свързване на биологично активното вещество с α-субединица на рецептора, рецепторът се променя от неактивно мономерно състояние в активно димерно състояние, при което два мономера се комбинират в мембранната равнина (виж фиг. 1.9). В този случай се стимулира ензимната активност на β-субединицата на рецептора на цитоплазмата, в резултат на това в целевата клетка се стартира каскада от биохимични реакции, които променят нейното функционално състояние.
Като трансмембранен ензим, образуващ рецептора, като правило се използват такива ензими като тирозин киназа или гуанилатциклаза.
Пример за рецептор на тирозин киназа са инсулиновите рецептори (виж Т. 1, стр. 435).
Пътят на предаване на сигнала от гуанилат циклаза започва с взаимодействието на а-субединицата на рецептора с ендогенно биологично активно вещество, например, с предсърден натриуретичен фактор (ANF), който е биологично активно вещество, секретирано от предсърдни клетки и участващо в регулирането на сърдечните контракции. В резултат на това взаимодействие настъпва промяна в конфигурацията на рецептора, която се състои в комбиниране на неговите мономери в димер. Този процес активира ензимната част на рецептора, разположена в неговата цитозолна β-субединица, т.е. ензимът гуанилат циклаза, който от своя страна насърчава повишаване на концентрацията на цикличния гуанидин-3,5 "монофосфат (cGMP) вторичен пратеник в целевата клетка. Увеличаването на концентрацията на cGMP в целевите клетки предизвиква каскада от биохимични реакции, които променят тяхното функционално състояние, например отпускане на гладкомускулните клетки съдове.
Рецепторите от тип III включват рецептори, които под въздействието на ендогенни биологично активни вещества - невротрансмитери, осигуряват преминаването на съответните йони през клетъчната мембрана, което води до промяна в неговия (мембранен) електрически заряд (потенциал).
В своята структура III tina рецепторите представляват канал, който прониква в липидния двуслой на клетъчната мембрана, образуван от няколко поли-тръбни единици (фиг. 1.10). Например, никотиновият (Н) рецептор е канал с диаметър 8 nm, образуван от пет полипептидни субединици (a - две, β, γ, d) (виж фиг. 1.10). Когато невротрансмитер ацетилхолин взаимодейства с част (домен), стърчаща над повърхността на клетъчната мембрана - α субединица на рецептора - структурата му се променя и се отваря централен канал, през който Na + йони навлизат в целевата клетка според градиента на концентрацията, което води до промяна във функционалната й функция дейност. В допълнение към Н-холинергичните рецептори, рецепторите за гама-аминомаслена киселина и възбуждащи аминокиселини принадлежат към рецептори тип III.
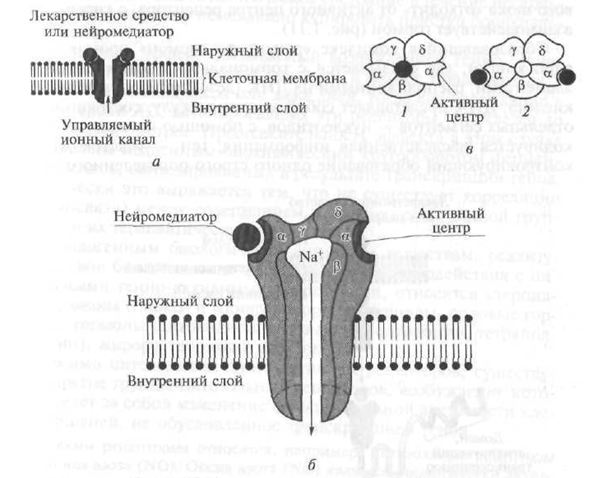
Фиг. 1.10. Структурната схема на рецептора тип 111:
a е схема; b - трансмембранен йонен канал (в контекста); в - трансмембранен йонен канал (изглед отгоре); / - канал в неактивно (затворено) състояние; 2 - канал в активно (отворено) състояние; а. β, γ, d - полипептидни субединици
Тип IV рецептори включват вътреклетъчни и ядрени рецептори. Биологично активните вещества, взаимодействащи с този тип рецептори, са липофилни (лесно разтворими в мазнини) съединения, поради което те лесно проникват в клетъчната мембрана и достигат до вътреклетъчните си рецептори. Вътреклетъчните рецептори включват рецептори за хормони, както и други биологично активни вещества.
Механизмът на взаимодействие на хормоните с вътреклетъчните рецептори е доста сложен, но той може да бъде представен схематично, както следва. По структура вътреклетъчният рецептор за хормони е полипептид, състоящ се от няколко функционални единици - домейни. При липса на хормон рецепторът е неактивен поради факта, че активният му център е блокиран от специализиран протеин - така наречения протеин от топлинен шок. В случай, че хормонът "се приближи" до рецептора, протеинът от топлинен шок "се отклонява" от активния център на рецептора, с който хормонът взаимодейства (фиг. 1.11).
Полученият рецептор-хормонен комплекс прониква в ядрото на клетката, където се свързва с чувствителни към хормона елементи, разположени на ДНК (дезоксирибонуклеинова киселина; ДНК е макромолекула, състояща се от отделни сегменти - нуклеотиди, с помощта на които наследствената информация се кодира в гените; генът - парче ДНК, което контролира образуването на един строго определен протеин
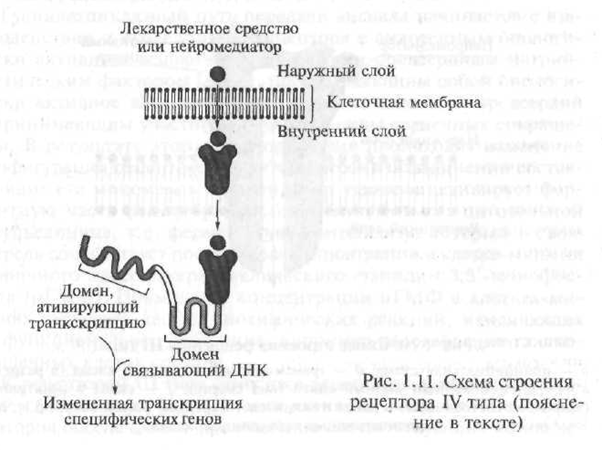
Фиг. 1.11. Схема на структурата на рецептор тип IV (обяснение в текста)
ка). В резултат на това взаимодействие започва процесът на генна транскрипция - процесът на прехвърляне на информацията, съдържаща се в генетичния код, от молекулата на ДНК в молекулата на информационната РНК (mRNA, syn: матрична РНК - мРНК). Транскрипцията е първата стъпка във формирането на протеини в клетка. Получената иРНК. напуска клетъчното ядро \u200b\u200bи се придвижва към рибозомите - вътреклетъчните органели, отговорни за синтеза на протеин в клетката. В специалната медицинска литература рецепторите, активирането на които причинява процеса на генна транскрипция, се означават като генетично активни рецептори.
По правило реакцията на целевите клетки на възбуждането на генно-активни рецептори се развива доста бавно, което е от много важно клинично значение.
Първо, реакцията на целевите клетки се забавя във времето, тъй като изисква синтеза на нови протеини, което обикновено отнема 20-30 минути, т.е. хормоните, активиращи рецептори тип IV, не са в състояние да променят патологичното състояние в рамките на няколко минути, например незабавно да спрат пристъп на бронхиална астма.
На второ място, ефектът, предизвикан от възбуждането на генно-активни рецептори, е доста дълъг и може да продължи няколко часа или дори дни, докато съдържанието в лекарствената система, активирала тези рецептори в кръвната плазма, намалява до нула много по-бързо. Продължителността на ефекта в този случай се дължи на бавната биохимична циркулация на ензими и протеини, синтезирани в резултат на генна транскрипция. Клинично това се изразява от факта, че няма връзка (връзка) между съдържанието на плазма в дадена група лекарства и техния терапевтичен ефект.
Ендогенните биологично активни вещества, които реализират своите биологични ефекти чрез взаимодействие с цитозолни генно-активни рецептори, включват стероидни хормони (глюко- и минералокортикостероиди, полови хормони), щитовидни хормони (трийодтиронин, тетрайодотиронин) и мастноразтворим витамин D.
В допълнение към цитозолните генно-активни рецептори, има и други групи цитозолни рецептори, чието възбуждане води до промяна във функционалната активност на целевите клетки, а не поради транскрипция на гени.
Такива рецептори включват, например, цитозолни рецептори за азотен оксид (N0). Азотният оксид (N0) е биологично активно вещество, образувано в съдовия ендотел. Като ендогенно биологично активно вещество, азотният оксид за първи път е изолиран от заешки пристанища от американския физиолог Р. Фърчготт през 1987 г. и е наречен „ендотелен релаксиращ фактор - ORF“. Азотният оксид е липофилно съединение, което лесно прониква в клетъчната мембрана, където взаимодейства със специфичните си цитозолни рецептори, което води до активиране на ензима гуанилат циклаза. Последният от своя страна стимулира синтеза на cGMP вторичния пратеник, който задейства каскада от вътреклетъчни биохимични реакции, водещи до релаксация на целевите клетки, съдовите гладко мускулни клетки.
Така понастоящем се разграничават четири основни механизма и съответно IV рецепторни типа, поради взаимодействието с които ендогенни биологично активни вещества и / или техните синтетични аналози, т.е. Лекарствата могат да повлияят на функционалното състояние на целевите клетки.
Това обаче не означава, че броят на известните рецептори за биологично активни вещества е ограничен до 4. Те са неизмеримо по-големи. Това се дължи на факта, че чрез един и същ основен механизъм на действие много голям брой ендогенни биологични вещества от различни химически структури могат да повлияят на функционалната активност на клетките. Например, невротрансмитерите норепинефрин и хистамин, които са различни по своята химична структура и следователно в рецепторите, с които взаимодействат, предават възбуждащ сигнал към целевите клетки по същия основен механизъм - стимулиране на активността на сигнални G протеини, т.е. и двете взаимодействат с рецептори тип I.
Следователно, всички известни понастоящем рецептори са класифицирани въз основа не само на характеристиките на предаване на сигнал към вътреклетъчните структури на целевите клетки, но и на имената на тези ендогенни биологично активни вещества, с които те конкретно си взаимодействат.
Трябва да се отбележи, че рецепторите са получили името си, като са взели предвид имената на ендогенни биологично активни вещества, с които те взаимодействат, много преди механизмите на предаване на сигнала към целевите клетки да станат известни.
Ендогенните биологично активни вещества, които реализират ефектите си чрез взаимодействие със специфичните им рецептори, включват невротрансмитери (ацетилхолин, норепинефрин, допамин, хистамин, серотонин и др.), Хормони, биологично активни вещества от тъканен произход - автокаиди (простагландини, тромбоксани, левкотриени и др.) брадикинин, ангиотензин и др.). В специалната медицинска литература всички тези вещества често се комбинират под термина „лиганди“ (от лат. Ligo - да се свързват, тоест вещество, способно да се свързва с рецептора).
Така рецепторите получават името си от името на техните специфични лиганди. Например рецепторите за допаминовия невротрансмитер се наричат \u200b\u200bдопамин, хормонът на инсулина - инсулин, левкотриен аугокаид - левкотриен и др.
В бъдеще в текста на учебника, за да се избегне объркване по отношение на механизма на предаване на сигнала от рецептора към вътреклетъчните формации, ще се използва терминът "рецепторен тип" и когато става въпрос за името на рецептора поради лиганда, взаимодействащ с него , ще се използва терминът "рецепторни видове".
По правило много рецептори от един и същи вид са разделени на няколко подтипа, например адренергичните рецептори са разделени на a- и β-адренергични рецептори, холинергичните рецептори на М- и N-холинергични рецептори и др. В повечето случаи подвидовете също се подразделят на по-малки групи: β1 - и β2-адренорецептори, N n - и Nm -холинергични рецептори и др.
Идентифицирането на подвида на рецепторите и изучаването на механизмите, по които ендогенните биологично активни вещества взаимодействат с тях е много важно за съвременната фармакология, тъй като позволява създаването на лекарства, които взаимодействат със строго определен подвид на рецептора. Така например, разделянето на β-адренергичните рецептори на β 1 (основно локализирано върху клетъчната мембрана на сърдечните клетки) и β 2 (локализирано например върху клетъчните мембрани на клетките на гладката мускулатура на бронхите) ни позволи да създадем лекарства, които селективно влияят на сърдечния мускул (β 1 -адреностимуланти ) - лекарството нонахлазин и селективно засяга гладката мускулатура на бронхите (β2-адреностимуланти) - лекарството салбутамол и др.
Трябва да се отбележи, че органите и тъканите на тялото не съдържат постоянен брой рецептори и / или техните подвидове, т.е. тя е променлива. Както патологичните процеси, така и лекарствата могат да променят броя на рецепторите в даден орган.
Например, коронарната болест на сърцето се придружава от увеличаване на броя на 3-адренергичните рецептори в сърдечния мускул, а при пациенти с хипертония се увеличава броят на а- и β-адренергичните рецептори.Дългосрочната употреба на антидепресант имипрамин намалява количеството на β-адренергичните рецептори в мозъчната тъкан. Има доста такива примери.
Афинитетът на ендогенни (произведени в тялото) невротрансмитери или лекарства за рецептори се характеризира с термина "афинитет", а скоростта и силата на свързването им с рецепторите се обозначават с термина "афинитет".
Естествено, взаимодействието на лекарствата с рецептора не е самоцел, но трябва да доведе до известна промяна в дейността на органи или тъкани на тялото.
Такава промяна или реакция, съответстваща на функционалното значение на този рецептор, се нарича вътрешна активност на лекарствата.
Лекарствата с присъща активност и рецепторен афинитет са агонисти, т.е. действат като ендогенни биологично активни вещества.
Например, фенилефрин, стимулатор на a-адренергичните рецептори, има ефект върху артериоли, подобни на невротрансмитер норепинефрин, т.е. той е агонист на a-адренергичните рецептори. В специалната медицинска литература, в допълнение към термина "агонист", понятието "рецепторен стимулант" или "миметик" понякога се използва, например, адреномиметик, т.е. Лекарства, които стимулират адренергичните рецептори.
Лекарствата, които имат рецепторен афинитет, но инхибират екзогенните и ендогенни агонисти да взаимодействат с рецептора, се наричат \u200b\u200bантагонисти.
Например блокерът на атропинов М-холинергичен рецептор пречи на взаимодействието на ацетилхолиновия невротрансмитер М-холинергичен рецептор; блокер (J-адренергични рецептори пропранолол, блокиращ β2-адреноструктури на белите дробове, предотвратява стимулиращия ефект на β2-адреностимулатора на салбутамол върху тях, т.е. атропин и пропранолол са антагонисти на съответните рецептори.
В специализираната медицинска литература, в допълнение към термина "антагонист", терминът "рецепторни блокери" или "литици", например, антихолинергици, т.е. лекарства, които блокират холинергичните рецептори.
Схематично взаимодействието на рецептори с агонисти и антагонисти е показано на фиг. 1.12.
Агонистите могат да упражняват както директни, така и косвени, т.е. косвени действия.

Фиг. 1.12. Схема на взаимодействие на рецептора с агониста (а) и антагониста (6) с рецептора (обяснение в текста)
Например, агонистът на опиоиден рецептор - лекарството морфин - осъзнава своите влияния чрез директно стимулиране на (p) p-, (капа) k- и (8) 8-опиоидни рецептори, т.е. има пряк стимулиращ ефект върху тях, докато симпатомиметичният ефедрин осъзнава клетъчните си ефекти чрез индиректно или косвено стимулиране на a- и β-адренергичните рецептори: помага за изместване на невротрансмитер норепинефрина от пресинаптични нервни окончания, инхибира неговото повторно поемане от нервните окончания и повишава чувствителността на a- и p- адренорецептори до норепинефрин и епинефрин и стимулира отделянето на адреналин от надбъбречната кора. По този начин ефедрин реализира фармакологичните си ефекти не директно, а чрез невротрансмитер, докато не взаимодейства директно с рецептора.
Антагонистите, както и рецепторните агонисти, могат да реализират своите фармакологични ефекти пряко или косвено, блокирайки съответните рецептори.
Например, блокерът на стомашния Н3 хистаминов рецептор, ранитидин, взаимодейства директно със стомашните Н2 хистаминови рецептори, инхибира базалната секреция на солна киселина, инхибира взаимодействието с ендогенния хистаминов невротрансмитер рецептор, т.е. директно блокира рецептора, докато симпатолитичният резерпин, влизайки в тялото, подобно на симпатомимистичния ефедрин „изхвърля“ невротрансмитера норепинефрин от пресинаптичните окончания, като едновременно блокира синтеза на нови „части“ норепинефрин, което води до бързо изчерпване на медиаторните резерви в пресинаптичните окончания. В резултат на това се създава състояние, когато а-адренергичните рецептори на съдовете "не работят" поради липсата на техния физиологичен стимулатор норадренапин. Можем да кажем, че симпатолитичният резерпин причинява функционална, индиректна блокада на a-адренергичните рецептори.
Някои лекарства комбинират свойствата на агонисти и антагонисти, т.е. при определени условия същите рецептори възбуждат или блокират. В случаите, когато стимулиращият компонент преобладава във фармакологичния ефект на лекарството, те казват, че лекарството е частичен или частичен агонист.
Механизмът на действие на частични или частични агонисти е, че тези лекарства, приближаващи се към рецептора, са фиксирани върху него. Поради особеностите на химическата им структура те го възбуждат по такъв начин, че да предизвикат не пълна, а само частична стимулираща реакция. В същото време те пречат на взаимодействието на други лекарства и / или съответния невротрансмитер с този рецептор.
Например, налорфинът, който се използва за лечение на предозиране на наркотични аналгетици, е частичен агонист. Налорфинът, „преминавайки“ към опиоидния рецептор, измества морфина от неговата връзка, тоест спира агонистичния ефект на морфина върху рецептора. Контактувайки с рецептора, налорфинът в малка степен има стимулиращ ефект върху него, който не е в състояние да потисне активността на дихателния център, т.е. има частичен агонистичен ефект върху опиоидния рецептор, паралелно с това го блокира за морфина, като по този начин проявява антагонистичен ефект по отношение на морфина. Недостатъкът на налорфина е, че в случай на предозиране, неговият стимулиращ ефект върху рецептора се засилва и той инхибира дихателния център, както и морфина.
В случаите, когато блокиращият ефект преобладава във фармакологичния ефект на лекарствата, те казват, че лекарството е антагонист със собствена или вътрешна активност.
Особености на механизма на действие на антагонистите със собствена активност могат да бъдат разгледани на примера на β 1 -адреноблокера на ацебуталол. Ацебуталолът принадлежи към групата на β1-адренергичните блокери със собствена вътрешна симпатомиметична активност, т.е. комбинира свойствата на блокер (литичен) и стимулант (миметик) на β1-адренорецептори.
Механизмът на действие на ацебуталол се дължи на особеностите на неговата химическа структура, поради които лекарството, от една страна, блокира миокардния β1 -адренорецептори, а от друга, стимулира тяхната активност, като по този начин симулира физиологичния ефект на медиаторите - катехоламините (виж Т. 1. p. 170).
Всички лекарства според особеностите на тяхното действие могат да бъдат разделени на две големи групи - със специфични и неспецифични ефекти.
Лекарствата, които имат неспецифичен ефект, включват лекарства, които имат широк спектър от фармакологична активност и различни точки на приложение на ефектите. Тази група включва витамини, биогенни стимуланти, антиоксиданти, адаптогени и др.
Лекарствата със специфичен ефект включват лекарства, характеризиращи се със свойствата на агонисти или антагонисти на съответните рецептори, т.е. основата на техния механизъм на действие е способността да взаимодействат със специфични за тях рецептори.
Трябва обаче да се отбележи, че фармакологичният ефект на лекарствата, които имат специфичен ефект върху всякакви рецептори, не винаги е силно специализиран, например намаляване на сърдечната честота (HR) или потискане на секрецията на стомашния сок. Общият фармакологичен ефект на много лекарства ще зависи от това колко и в кои органи и тъкани на тялото са локализирани специфични рецепторни образувания за това лекарство.
Например, β-адренергичните рецептори са разположени в сърдечния мускул, съдовата стена, бронхите, матката, мастната тъкан, скелетния мускул и др. В резултат на това лекарствата, които стимулират β-адренергичните рецептори, до известна степен ще предизвикат увеличаване на силата и честотата на сърдечните контракции, разширяване на кръвоносните съдове, бронхите и по-ниския тонус на матката, т.е. имат системен ефект върху организма.
Специфичният ефект на лекарствата може да бъде селективен или селективен и съответно неселективен или неселективен. Селективността на лекарствата се определя от това дали засяга всички подвидове на някакви рецептори или техните специфични подвидове.
Например, неселективният β-адреноблокер пропранолол блокира както β1-адренорецепторите, разположени в миокардната тъкан, така и β2-адренорецепторите, разположени по-специално в белодробната тъкан, т.е. има неселективен (неселективен) ефект върху двата подвида на β-адренергични рецептори , като има предвид, че селективният β 1 -адренергичен блокер атенолол в терапевтични дози блокира само β 1 миокардни адренергични рецептори, т.е. упражнява селективен (селективен) ефект върху специфичен β-адренергичен рецептор.
Изясняване на физиологичната роля и локализацията на рецепторните подвидове позволява създаването на високоефективни лекарства, които селективно действат на различни подвидове рецептори.
Например, лекарствата, които блокират β1-адренергичните рецептори, се използват широко в клиниката за лечение на коронарна болест на сърцето, артериална хипертония и лекарства, които стимулират β2-адренергичните рецептори, намериха своето приложение за лечение на бронхиална астма. Лекарствата, които блокират Н1 хистаминовите рецептори, за предпочитане се предписват в клиничната практика за предотвратяване и / или спиране алергични реакциидокато лекарствата, блокиращи Н2 хистаминовите рецептори, са ефективни при лечение на стомашни и дванадесетопръстни язви.
Създаването на лекарства със селективен ефект до голяма степен оптимизира клиничната им употреба, тъй като те значително намаляват страничен ефект.
Основата на механизма на действие на лекарствата като правило е способността им да инициират (задействат) сложни биохимични и / или биофизични процеси, които в крайна сметка променят и / или оптимизират функционалната активност на целевата клетка. Лекарствата могат да реализират ефекта си върху органите и / или целеви клетки чрез: директно химично взаимодействие; физико-химично взаимодействие върху клетъчната мембрана; действия върху специализирани ензими; действия върху регулаторните гени; действия върху специфични рецептори.
Директно химическо взаимодействие на лекарствата.
Този механизъм на действие на лекарствата е доста рядък и може да се реализира извън клетката, например в лумена на стомаха или червата. Същността му се състои във факта, че лекарствата влизат в пряка химическа реакция с молекули и / или йони, които се образуват в организма при нормално или патологично състояние. Пример за пряко химическо взаимодействие е химическата реакция на неутрализиране на солна киселина на стомаха при прием на антиацидни лекарства.
Физико-химично взаимодействие на лекарства върху клетъчната мембрана.
Една от основните функции на цитоплазмената мембрана е осъществяването на йонен обмен между цитоплазмата и извънклетъчната среда. Трансмембранният йонен обмен може да се извърши и чрез специални трансмембранни йонни канали, зависими от напрежението - натрий, калий, калций, хлор и др. Някои лекарства, достигайки клетъчната мембрана, взаимодействат с тези канали и променят функционалната им активност. Така например, антиаритмичният ефект на лекарството от клас IA - хинидин - се основава на способността му да блокира преминаването на Na + йони през трансмембранни натриеви канали.
Ефектът на лекарствата върху специализираните ензими.
Сравнително малко количество лекарства реализира фармакологичния му ефект чрез промяна на активността на някои специализирани клетъчни ензими. Наричат \u200b\u200bсе лекарства, които повишават активността на клетъчните ензими индукториензими. Такова действие се притежава например от хапчета за сън и антиконвулсантно лекарство фенобарбитал, което значително засилва активността на микрозомалните чернодробни ензими. Биологичното значение на този ефект на фенобарбитала и свързаните с него лекарства ще бъде разгледано по-долу.
Наричат \u200b\u200bсе лекарства, които инхибират активността на специализирани ензими инхибиториензими. Така например, антидепресант от групата на инхибиторите на моноаминооксидазата (МАО), лекарството пирлиндол реализира антидепресантния си ефект чрез инхибиране на активността на ензима МАО в централната нервна система. Способността да инхибира активността на ензима ацетилхолинестераза е в основата на фармакологичната активност на антихолинестеразните лекарства, например физостигмин. Известно е, че при физиологични условия ацетилхолинестеразата инактивира (унищожава) ацетилхолин, невротрансмитер, който предава възбуждане в синапсите на парасимпатиковата нервна система. Физостигминът, потискащ активността на ацетилхолинестеразата, насърчава натрупването в синапсите на парасимпатиковата система на невротрансмитера ацетилхолин, в резултат на което се повишава тонусът на парасимпатиковата нервна система, което се проявява на системно ниво чрез развитието на брадикардия, понижаване на кръвното налягане (на гастроинтетичния тракт), на стомашно-чревния тракт (понижаващо кръвно налягане) ученик и т.н. Лекарствата могат да взаимодействат обратимо и необратимо с ензимите. Например, лекарството еналаприл обратимо инхибира активността на ангиотензин-конвертиращия ензим, което в частност води до понижаване на кръвното налягане, докато токсичните вещества от органофосфор необратимо инхибират активността на ацетилхолинестеразата. Ефектът на лекарствата върху регулаторните гени.В момента учените правят опити да създадат лекарства, които реализират фармакологичните им ефекти, като влияят пряко върху физиологичната активност на регулаторните гени. Тази тенденция изглежда особено обещаваща, след като структурата на човешкия геном беше дешифрирана през 2000 г. Смята се, че селективното нормализиране на функцията на регулаторните гени под въздействието на лекарства ще направи възможно постигането на успех в лечението на много заболявания, включително преди това неизлечими. Ефектът на лекарствата върху рецепторите.Преди да преминем към характеристиките на взаимодействието на лекарствата с рецепторите, е необходимо да се изясни какво имаме предвид под термина „рецептор“ (от лат. recipio - вземам, вземам). От курса на физиологията е известно, че терминът "рецептор" означава високоспециализирани формации, които са в състояние да възприемат, трансформират и предават енергията на външен сигнал към нервната система. Тези рецептори се наричат сетивен(от лат. sensus - усещане, усещане, възприятие). Сензорните рецептори включват рецептори на органите на слуха, зрението, обонянието, вкуса, допира и т.н. Сензорните рецептори на тези органи принадлежат към така наречените екстерорецептори. Ако наличието на сетивни органи, които реагират на външни стимули на дразнене, е било известно от древни времена, то присъствието на сензорни рецептори вътре в тялото е било под въпрос до средата на 19 век. За първи път руският физиолог I.F. Цион, който през 1866 г. показа спад на кръвното налягане поради дразнене на аортата при експеримент със заек. Това откритие породи търсенето и изучаването на рецептори, разположени вътре в тялото, и самите тези рецептори бяха наречени interoрецептори. Kначалото на XX век беше разкрит достатъчен брой сензорни интерорецептори и е доказана тяхната важна роля в регулацията на физиологичните функции на организма. През 1905 г. J. Langley доказва, че когато лекарството се прилага върху клетъчната мембрана, се развива фармакологичен ефект, ако се прилага само върху конкретна област от него. Освен това този сайт представлява само малка част от общата площ на клетъчната повърхност. Това наблюдение позволи на J. Langley да заключи, че на клетъчната мембрана съществуват специализирани рецепторни сайтове, взаимодействащи с лекарства. Въпреки това приоритетът при създаването на рецепторната теория за действието на лекарствата принадлежи на немския физиолог П. Ерлих, който през 1906 г. въвежда понятието „рецептор“ и формулира постулата „лекарството не работи, ако не се фиксира върху клетъчната мембрана“. Според теорията на P. Ehrlich, молекулата на лекарството има две функционално активни групи, едната от които осигурява фиксацията му върху клетъчната повърхност в областта на лекарствения рецептор, а втората функционална група взаимодейства с рецептора и задейства сложна верига от биохимични реакции, които променят неговата (клетъчна) физиологична активност , По този начин още в началото на 20 век. стана ясно, че има поне два класа интерорецептори: сензорни рецепторипредаване на информация за състоянието на вътрешните органи и телесните тъкани в централната нервна система; клетъчни рецепторикоито взаимодействат с лекарства, които променят функционалната активност на целевите клетки.<>Веднага трябва да се отбележи, че в бъдеще в текста на учебника, за да се избегне объркване в терминологията, рецепторите за лекарства и биологично активни вещества, т.е. клетъчен или циторецептори,ще бъдат обозначени с термина "рецептор", докато сензорните интерорецептори ще бъдат обозначени с термин, характеризиращ тяхната функционална активност, например "барорецептори", "рецептори на болка" и т.н. Откритието от П. Ерлих върху клетъчната мембрана на лекарствените рецептори послужи като отправна точка за развитието на фармакологичната наука, по-специално фармакодинамиката, една от основните задачи на която е да се проучат рецепторните механизми на действие на лекарствата. Понастоящем са разкрити структурата на голям брой клетъчни рецептори, особеностите на взаимодействието на определени биологично активни съединения с тях, което даде възможност, от една страна, да се разбере механизмът на действие на известни лекарства, а от друга, беше основата за създаването на нови високоефективни лекарства. Естествено е трудно да си представим, че в хода на еволюцията в човешкото тяло се образуват рецептори за различни синтетични (химически получени) лекарства, особено след като огромното мнозинство лекарства, представени на съвременния фармацевтичен пазар, са синтезирани през последните 50 години или по-малко. Доказано е, че рецепторният апарат на клетката е много древна функционално-структурна формация. И така, a- и b-адренорецепторите (рецептори, взаимодействието на които норепинефрин и адреналин влияят върху функционалната активност на клетката) се намират не само в клетките на животните, но и в клетъчните мембрани на растителните клетки, например в клетките на растителната нителла, където a- и b- адренорецепторите регулират движението на протоплазма (съдържанието на клетките). Тогава какви са рецепторите за лекарства, открити от П. Ерлих, и защо те взаимодействат с тях? В момента няма съмнение, че така наречените лекарствени рецептори всъщност са рецептори за ендогенни (произвеждани в организма) биологично активни вещества, участващи в регулирането на функционалната дейност на вътрешните органи и телесните тъкани. Такива биологично активни съединения включват вещества, освободени от нервните окончания в момента на предаване на нервния сигнал, както и хормони, витамини, аминокиселини и др. За всяко ендогенно биологично активно вещество има строго специфични рецептори за него. Така например, биологично активното вещество, произведено в организма, адреналинът, може да активира строго специфични а - и b-адренергичните рецептори, а глюкокортикостероидите - хормони на надбъбречната кора - взаимодействат само със строго специфични за тях глюкокортикостероидни рецептори. Синтетичните лекарства, които реализират ефектите си, взаимодействайки с рецепторния апарат на клетката, по своята химическа структура, са повече или по-малко подобни на ендогенни биологично активни съединения, които взаимодействат с подобни рецептори. Например синтетичните вазоконстрикторни (причиняващи вазоконстрикция) лекарства фенилефрин са близки по своята химическа структура с ендогенното биологично активно вещество норепинефрин, следователно, подобно на норепинефрин, той има способността да стимулира a-адренорецепторите. Понякога, поради особеностите на тяхната химическа структура, лекарствата могат да взаимодействат не със самия рецептор, а със съседната част на клетъчната мембрана. Тъй като в този случай лекарството не взаимодейства със самия рецептор, а със съседната част на клетъчната мембрана, те говорят не за възбуждащ или блокиращ ефект върху рецептора, а allosteметрично(от гръцки allos - друг, различен) ефект, или ефект. В резултат на това може да настъпи промяна както в структурата на мембраната, съседна на рецептора, така и в отделните компоненти на самия рецептор, което може да доведе до промяна в чувствителността на рецептора към биологично активно вещество, специфично за него. В случаите, когато чувствителността на рецептора към биологично активно вещество се увеличава, говорете сенсибилизация(от лат. sensus - усещане) или около сенсибилизация(от лат. sensibilis - чувствителност) на рецептора и в тези случаи, когато чувствителността на рецептора намалява, говорете за десенсибилизациярецептор. Особеността на алостеричния ефект се крие във факта, че лекарствата, които имат този вид механизъм на действие, не влияят пряко върху предаването на нервен импулс, а го променят в желаната посока. Например, механизмът на действие на анксиолитиците (лекарства против тревожност; синоними: транквилизатори), по своята химическа структура са производни на бензодиазепин, се основава на феномена на алостерично възбуждане на постсинаптичните бензодиазепинови рецептори. Възбуждането на последните от своя страна насърчава активирането на инхибиторните постсинаптични рецептори на гама-аминомаслена киселина (GABA рецептори), което се проявява клинично чрез елиминиране на симптоми на невротични заболявания като тревожност, тревожност, страх и др. Рецепторите, взаимодействащи с които, биологично активно вещество или лекарство по някакъв начин променя функционалното състояние на целевата клетка, се наричат специфични. В допълнение към специфичните рецептори, т.нар неспецифичниза лекарствени рецептори. В специализираната медицинска литература тези рецептори се наричат \u200b\u200bоще „мястото на загубата“ на лекарствата. Свързвайки се с такива рецептори, лекарствата нямат никакъв биологичен ефект, но самите те стават биологично неактивни. Пример за този тип рецептори могат да служат като рецептори, разположени върху плазмените протеини, по-специално върху водоразтворимите протеини - албумин. Структурата на рецепторите е доста сложна, но повечето от тях са протеинови макромолекули или гликопротеини, които също могат да включват йони, липиди, нуклеинови киселини и др. Рецептор, т.е. образуващата го протеинова макромолекула се характеризира със специфично, специфично за всеки рецептор, пространствено разположение на неговите химични групи. Протеиновата макромолекула, образуваща рецептора, може да бъде интегрирана (потопена) в липидната двуслойна на цитоплазмената мембрана или локализирана вътре в клетката. Основната функция на клетъчния рецептор е да „разпознава“ химичен сигнал, предаван към него чрез ендогенно биологично активно вещество и / или лекарства и да го трансформира в съответния биохимичен и / или биофизичен клетъчен отговор. По-рано се смяташе, че лекарствата или ендогенните биологично активни вещества взаимодействат с рецепторите от типа „ключ и заключване“, т.е. рецепторът има такава структура, която позволява на лекарството да намери "вашия" рецептор, да се свърже с него и, както би било, "включете" и "изключете". С развитието на медицинската наука обаче стана очевидно, че това не е напълно вярно. Понастоящем молекулните процеси на превръщане на извънклетъчните сигнали във вътреклетъчна, регулираща клетъчната функция, вече са проучени доста добре. механизми, които водят до ефекта от взаимодействието на ендогенни биологично активни вещества или лекарства с рецептори. При взаимодействие с рецептора на ендогенно биологично активно вещество и / или активно лекарство като него структура- пространствена промяна под формата на протеинова макромолекула, която е спусъкът за различни вътреклетъчни процеси, които определят отговора на целева клетка към медиатор и / или лекарство. Например, активирането на b2-адренорецептори на гладката мускулатура на бронхите под въздействието на b2-адреностимулаторен фенотерол води до повишаване на активността на ензима аденилатциклаза, което допринася за натрупването на цикличен аденозинов монофосфат (cAMP) в клетката и в резултат на това клетъчната релаксация. Като цяло биологичните термини, клетъчните рецептори могат да се считат за строго специализирани „сетивни органи“ на клетките, чрез които те възприемат „информация“, излъчвана например от централната нервна система и / или ендокринната система. Въпреки важната роля на рецепторния апарат, рецепторите заемат само незначителна част от клетъчната мембрана. Например, М-холинергичният рецепторен апарат на клетка заема не повече от 1/6 000 от нейната повърхност. Изучаването на характеристиките на взаимодействието на лекарствата с рецептора, от една страна, ни позволява да разберем основата на молекулярния механизъм на неговото действие, а от друга, дава информация какви промени трябва да се направят в структурата на лекарствата, за да се повиши способността му да взаимодейства с този рецептор, т.е. позволява целеви синтез на нови високоефективни лекарства. При физиологични условия различните клетъчни рецептори не функционират независимо, но са в постоянно взаимодействие помежду си, като по този начин регулират специфичната активност на клетката. Например, активирането на сърдечните b-адренергични рецептори от ендогенни норепинефрин причинява по-специално увеличаване на броя на сърдечните контракции и активирането на М-холинергичните рецептори на сърдечните клетки от ендогенния ацетилхолин, напротив, причинява намаляване на броя на сърдечните контракции. Голям принос за разбирането на рецепторните механизми на действие на лекарствата направиха откриването на пре- и постсинаптични рецептори. Bзаспивания(от гръцки резюме - връзка) е специализирана контактна зона между нервните клетки или други възбудими структури на тялото, която осигурява предаването на входяща информация и запазването на нейното информационно значение. Изучаването на структурата и функционалната роля на синапсите започва в края на 19 век. след като испанският хистолог С. Рамон-и-Каджал (S. Ramon at Cajal) предложи наличието на специализирана предавателна система в централната нервна система. Синапсите получават името си през 1897 г., когато английският физиолог Ч. Шеррингтън предлага този термин да се отнася до зоната на контакт между нервните клетки. Понастоящем има три типа синапси: 1) "електрически" синапси, при които информацията се предава чрез прехвърляне на електрически сигнал от пресинаптична мембрана. Този тип синапс се нарича efaps(от гръцки ephapsis - тесен контакт); 2) "химически" синапси, при които информацията се предава чрез специални биологично активни вещества - невротрансмитери(от гръцки неврон - нерв и лат. посредник - посредник; синоними: медиатор); 3) „смесени“ синапси, при които информацията се предава както химически, така и електрически. Фармакологичните ефекти на огромното мнозинство лекарства, които влияят върху функциите на синапсите, се реализират чрез ефекта им върху определен етап на предаване на сигнала в химичните синапси, т.е. в синапси от втория вид. По правило химичните синапси се класифицират по невротрансмитери, които предават нервни импулси, както следва: синапсите, в които ацетилхолинът действа като медиатор, се наричат \u200b\u200bхолинергични; синапси, при които адреналинът и норепинефринът действат като медиатор се наричат \u200b\u200bадренергични; синапси, в които АТФ и аденозинът действат като медиатор се наричат \u200b\u200bпуринергични; синапси, в които гама-аминомаслената киселина действа като медиатор, се наричат \u200b\u200bGABA-ергични и др. В момента структурата на синапса е добре разбрана. Синапсът се състои от пресинаптичен процес на нервна клетка (аксонов край) и "сигнал" -приемащ апарат, разположен върху мембраната на ефекторна ("изпълнителна") клетка.
Фармакодинамика - фармакологични ефекти, механизми на действие, локализация на действието, видове действие на лекарствата.
Фармакологични ефекти лекарствено вещество - промени в дейността на органите, телесните системи, които веществото причинява (например засилени сърдечни контракции, понижаване на кръвното налягане, стимулиране на умствената дейност, премахване на страх и напрежение и др.).
Всяко вещество предизвиква редица фармакологични ефекти, характерни за него. Във всеки случай се използват само определени ефекти на лекарственото вещество, които са определени като главното ефекти. Останалите (неизползвани, нежелани) фармакологични ефекти се наричат вреден.
Механизми на действие лекарства - начините, по които веществата причиняват фармакологични ефекти са много разнообразни. Основните възможности за механизмите на действие включват действия по:
- специфични рецептори;
- ензими;
- йонни канали;
- транспортни системи.
Повечето лекарства действат върху специфични рецептори. Тези рецептори са най-често представени от функционално активни протеинови молекули, взаимодействието с които поражда биохимични реакции, които водят до фармакологични ефекти.
Има специфични рецептори, свързани с клетъчните мембрани (мембрана) и вътреклетъчни рецептори (цитоплазмена, ядрена).
Мембранните рецептори (рецептори на цитоплазмената мембрана) се делят на:
- рецептори, директно свързани с йонни канали;
- рецептори, директно конюгирани с ензими;
- рецептори, които взаимодействат с G-протеини.
K рецептори, директно свързани с йонни канали, включват по-специално Ν-холинергични рецептори и GABA A-рецептори.
Когато се стимулират Ν-холинергичните рецептори (чувствителни към никотин холинергични рецептори), натриевите канали, директно свързани с тях, се отварят. Стимулирането на Ν-холинергичните рецептори води до откриване на Na + канали, навлизане на Na + йони в клетката, деполяризация на клетъчната мембрана и вълнуващ ефект.
GABA A рецепторите са директно свързани с хлоровите канали. Стимулирането на GABA рецепторите води до откриване на Cl - канали, навлизане на Cl - йони, хиперполяризация на клетъчната мембрана и инхибиторен ефект.
K рецептори, които са пряко свързани с ензимитевключват, по-специално, инсулинови рецептори, директно конюгирани с тирозин киназа.
G-протеинови рецептори - М-холинергични рецептори (чувствителни към мускарините холинергични рецептори), адренергични рецептори, допаминови рецептори, опиоидни рецептори и др.
G-протеините, т.е. GTP-свързващи протеини, са локализирани в клетъчната мембрана и се състоят от α-, β- и γ-субединици. Когато лекарството взаимодейства с рецептор, α-субединицата на G-протеина се свързва с GTP (GTP) и действа върху ензими или йонни канали.
Един рецептор взаимодейства с няколко G-протеини и всеки комплекс от α-субединица на G-протеин с GTP действа върху няколко ензимни молекули или върху няколко йонни канала. По този начин се осъществява механизмът на усилвател (усилвател): когато се активира един рецептор, активността на много ензимни молекули или много йонни канали се променя.
Един от първите открити бяха G-протеини, свързани с β1-адренорецептори на сърцето. Когато се активира симпатичната инервация на сърцето, β1 -адренорецепторите се възбуждат; аденилатциклазата се активира чрез G-протеини; сАМФ се образува от АТФ, активира се протеин киназата, под действието на която Са 2+ каналите се фосфорилират и отварят.
Увеличаването на входа на Ca 2+ йони в клетките на синоатриалния възел ускорява четвъртата фаза на потенциала на действие, честотата на генерираните импулси се увеличава - сърдечните контракции стават по-чести.
Откриването на Ca 2+ канали във влакната на работещия миокард води до повишаване на концентрацията на Ca 2+ в цитоплазмата (Ca 2+ входът насърчава освобождаването на Ca 2+ от саркоплазмения ретикулум). Ca 2+ йони се свързват с тропонин С (компонент на тропонин-тропомиозин); По този начин инхибиторният ефект на тропонин-тропомиозин върху взаимодействието на актин и миозин намалява - сърдечните контракции се засилват (фиг. 10).
Фиг. 10. Механизмът за увеличаване и засилване на сърдечните контракции по време на стимулация на β1 -адренорецепторите. променлив ток - аденилатциклаза; PC - протеин киназа; CA - синоатриален възел; ТТМ - тропонин-тропомиозин.
Когато се активира парасимпатиковата инервация на сърцето (вагусните нерви), М2 -холинергичните рецептори се възбуждат и чрез G-протеините аденилатциклазата се инхибира - сърдечните контракции се отслабват и отслабват (главно предсърдното контракция се отслабва, тъй като парасимпатиковата инервация на вентрикулите е сравнително лоша).
По този начин, G-протеините могат да имат стимулиращ и инхибиращ ефект върху аденилатциклазата. Стимулиращите G-протеини са обозначени като G s-протеини (стимулират) и инхибиторни G-протеини (G-протеини (инхибират) (фиг. 11).
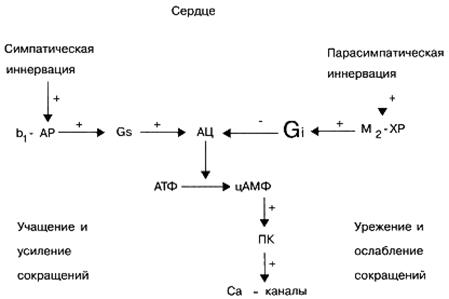
Фиг. 11. Механизмът на промяна в честотата и силата на сърдечните контракции по време на стимулация на симпатиковата и парасимпатиковата инервация.
Холеровият токсин активира Cs протеините (това води до активиране на аденилат циклазата, а при холерата се проявява чрез секреция на течност през чревния епител).
Коклюшният токсин активира G i протеините.
Когато М1 -холинергичните рецептори, МЗ -холинергичните рецептори, α-адренорецепторите се възбуждат чрез протеини G q, фосфолипаза С се активира, което допринася за образуването на инозитол-1,4,5-трифосфат и диацилглицерол от фосфатидилинозитол-4,5-дифосфат ,
Инозитол-1,4,5-трифосфат действа върху рецепторите на чувствителната към него саркоплазмена ретикулумна мембрана и стимулира освобождаването на Са 2+ йони от саркоплазмения ретикулум (фиг. 12). При стимулирането на α-адренорецепторите на кръвоносните съдове това води до свиване на гладката мускулатура на съдовете и стесняване на съдовете (фиг. 13).

Фиг. 12. Ефектът на фосфолипаза С върху нивото на цитоплазмата C a2 +.
Чувствителността на рецепторите към агонистите и броят на рецепторите постоянно се променят. И така, след стимулиране на β 1 -адренорецепторите от агонист на β1-адренорецепторите, фосфорилатът чрез специална рецепторна киназа се свързва с протеин на β-арестилин и в този комплекс губи способността си да взаимодейства с G-протеини (десенсибилизация на рецепторите). Комплексът от β1 -адренорецептори с β-арестилин се абсорбира от клетката чрез ендоцитоза (интернализация на рецепторите) и се улавя от ендозоми и лизозоми. В ендозомите молекулите на β1-щама се отделят от рецепторите, които се интегрират отново в клетъчната мембрана; възстановява се чувствителността на рецептора към агонистите (рецепторна ресенсибилизация). В лизозомите се случва разрушаването на рецепторните молекули (надолу-регулация) (фиг. 14).

Фиг. 13. Механизмът на свиване на гладките мускули на кръвоносните съдове по време на стимулация на симпатичната инервация. FLS - фосфолипаза С; FIF 2 - фосфатидилинозитол-4,5-дифосфат; Ако 3 - инозитол-1,4,5-трифосфат; CP - саркоплазмен ретикулум; MLCK - киназа от леки вериги на миозин.

Фиг. 14. Десенсибилизация и понижаваща регулация на β-адренергичните рецептори.
K вътреклетъчни рецептори включени са кортикостероидните и половите хормонални рецептори. По-специално, глюкокортикоидните рецептори са локализирани в цитоплазмата на клетките. След като глюкокортикоидът се комбинира с цитоплазмени рецептори, глюкокортикоидният рецепторен комплекс прониква в ядрото и влияе върху експресията на различни гени.
Способността на веществата да се свързват с рецепторите (склонността на веществата да се свързват с рецепторите) е обозначена с термина „ афинитет". По отношение на едни и същи рецептори афинитетът на различните вещества може да бъде различен. За да характеризирате афинитета, използвайте индикатора рK D е отрицателният логаритъм на дисоциационната константа, тоест концентрацията на веществото, при която са заети 50% от рецепторите.
Вътрешна дейност - способността на веществата да стимулират рецепторите; определено от фармакологичен ефектсвързани с активиране на рецепторите.
При нормални условия няма пряка връзка между афинитета и вътрешната активност. Веществото може да заеме всички рецептори и да причини слаб ефект, и обратно, веществото може да заеме 10% от рецепторите и да причини максимален ефект за тази система.
агонисти - вещества с афинитет и вътрешна активност.
Пълни агонисти притежават афинитет и максимална вътрешна активност (те могат да предизвикат максимален ефект за дадена система), дори ако заемат част от специфични рецептори.
Частични (частични) агонисти притежават афинитет и по-малко от максимална вътрешна активност (те могат да причинят само по-малко от максималните ефекти, дори ако заемат 100% от специфичните рецептори).
антагонистипритежават афинитет, но не притежават вътрешна активност и пречат на действието на пълни или частични агонисти (изместват агонистите от комуникацията с рецепторите).
Ако действието на антагониста се елиминира чрез увеличаване на дозата на агониста, такъв антагонизъм се нарича конкурентен.
Частичните агонисти могат да бъдат антагонисти на пълни агонисти. При липса на пълен агонист, частичен агонист стимулира рецепторите и предизвиква слаб ефект. Когато взаимодейства с пълен агонист, частичен агонист заема рецептори и инхибира действието на пълен агонист. В този случай ефектът на пълен агонист е отслабен.
Например, пиндолол, частичен агонист на β-адренорецепторите, причинява слаба тахикардия при липса на ефекти на симпатична инервация върху сърцето. Но с повишаване на тона на симпатичната инервация, пиндололът действа като истински β-блокер и причинява брадикардия. Това се дължи на факта, че частичният агонист пиндолол отслабва ефекта на медиатора норепинефрин, който е пълен агонист по отношение на β-адренергичните рецептори на сърцето.
Агонисти на антагонисти - вещества, които действат различно на подтипове на едни и същи рецептори: стимулират някои рецепторни подтипове, а блокират други. Например наркотичният аналгетик налбуфин има различни ефекти върху подтипове на опиоидни рецептори. Налбуфин стимулира κ рецепторите (и следователно намалява чувствителността към болка), а µ рецепторите блокира (и следователно е по-малко опасен от гледна точка на наркотичната зависимост).
Пример за въздействието на веществата върху ензими може да има действие на антихолинестеразни средства, които блокират ацетилхолинестеразата (ензим, който разгражда ацетилхолина) и по този начин усилва и удължава действието на ацетилхолина.
Известни лекарствени вещества, които стимулират или блокират йонни канали клетъчни мембрани, тоест канали, които селективно провеждат йони Na \u200b\u200b+, K +, Ca 2+ (натриеви, калиеви, калциеви канали) и др. Например:
Местните анестетици блокират Na + каналите;
Антиаритмичните лекарства от клас I (хинидин, лидокаин) блокират Na + каналите;
Миноксидил активира К + каналите;
Хипогликемичните агенти от групата на производни на сулфонилуреята блокират АТФ-зависимите К + канали;
Верапамил, нифедипин блокира Ca 2+ каналите.
Пример за въздействието на веществата върху транспортни системи може да има действие:
Резерпин (блокира везикуларното поглъщане на допамин и норепинефрин);
Сърдечни гликозиди (инхибират Na + / K + -ATPase);
Трициклични антидепресанти (блокират обратното поемане на невроните на норепинефрин и серотонин);
Блокатори на протонната помпа (омепразол и др.).
Възможни са и други механизми на действие. Например диуретичният манитол увеличава диурезата чрез повишаване на осмотичното налягане в бъбречните канали. Антиатеросклеротичният агент - колестипол - свързва (секвестрира) жлъчните киселини, предотвратява абсорбцията им в червата, поради което се активира образуването на жлъчни киселини от холестерола в черния дроб и нивото на холестерола в хепатоцитите намалява.
Механизмите на действие на различни лекарства са изучени в различна степен. В процеса на изучаването им идеите за механизмите на действие могат не само да станат по-сложни, но и да се променят значително.
Концепцията за „ локализация на действието„Означава основното (ите) място (и) на действие на някои лекарствени вещества. Например, сърдечните гликозиди действат главно върху сърцето.
Към концепцията за „ видове действия»Относно местните и общи (резорбтивни) действия, рефлекторно действие, първично и вторично действие, пряко и косвено действие.
Пример за локално действие може да бъде действието на локални анестетици.
Повечето лекарства имат общ (резорбтивен) ефект, който обикновено се развива след абсорбция (резорбция) на вещество в кръвта и разпределението му в организма.
Както с локални, така и с резорбтивни действия, веществата могат да възбуждат различни чувствителни рецептори и да предизвикат рефлекторни реакции.
Основният ефект на лекарственото вещество са неговите ефекти, които се използват във всеки отделен случай. Всички останали ефекти се оценяват като прояви на странични ефекти.
Лечебните вещества могат да упражняват определени органи директно действие, Освен това ефектът от лекарствата може да бъде косвен. Например, сърдечните гликозиди имат пряк ефект върху сърцето, но, подобрявайки функционирането на сърцето, тези вещества увеличават кръвоснабдяването и функциите на други органи (косвен ефект).